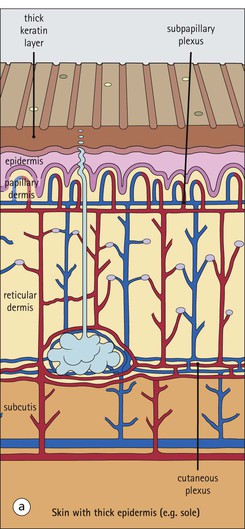
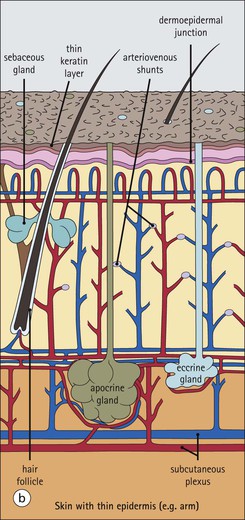
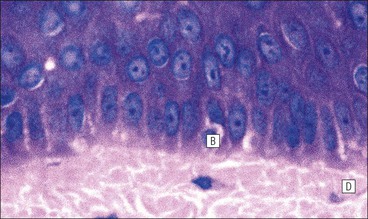
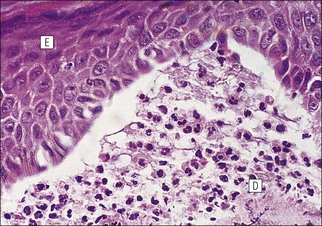
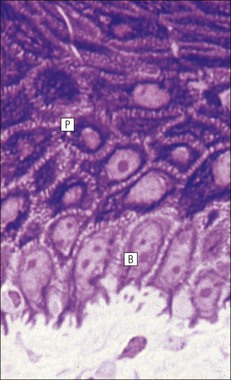
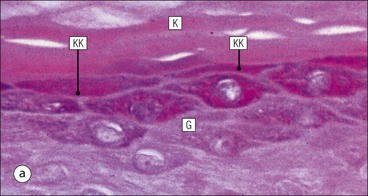
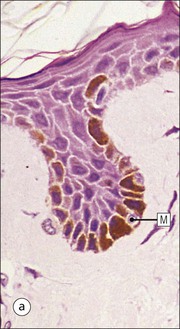
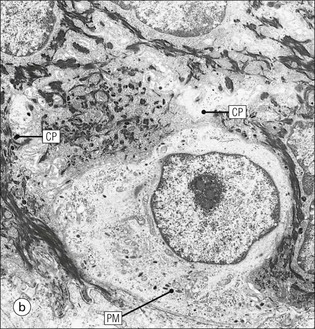
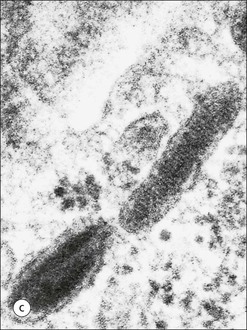
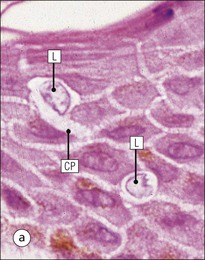
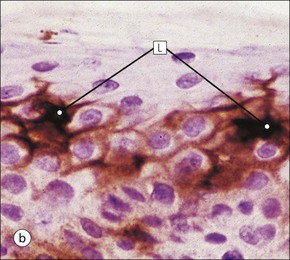
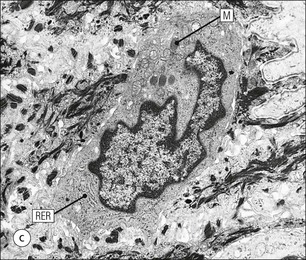
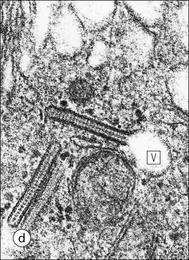
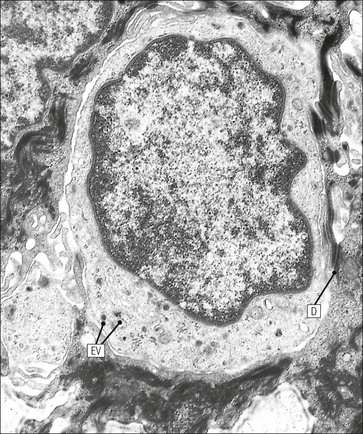
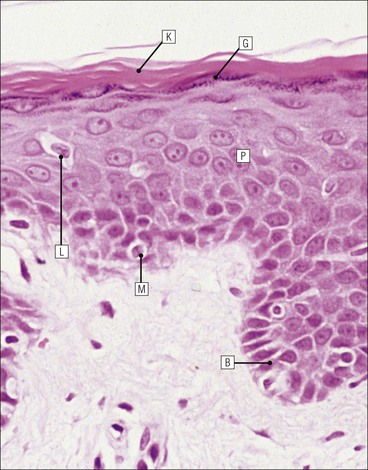
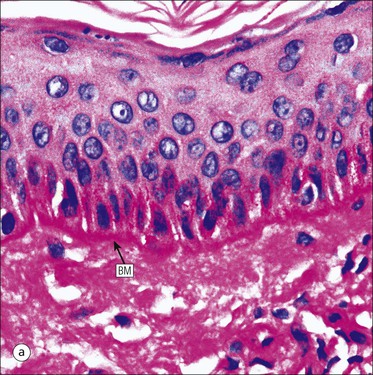
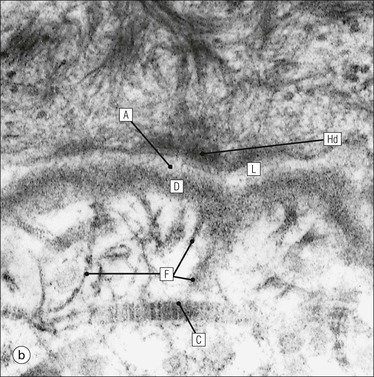
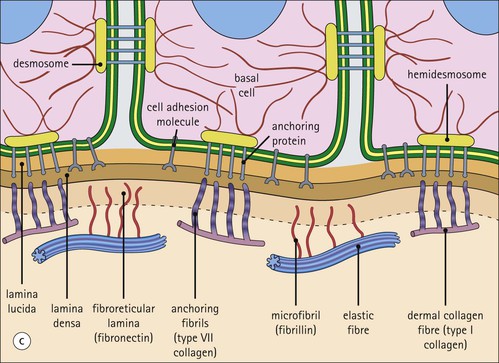
The skin is an extensive organ covering the exterior of the body. It varies in structure from site to site according to specific functions, which include: The breast is a highly modified area of skin with specialized sweat glands to produce nutritious secretions under hormonal influences. The skin is composed of two main layers: the epidermis and dermis, and a variable third layer, the subcutis. The epidermis is the surface epithelial layer in contact with the external environment. Downgrowths of this layer produce sweat glands, hair follicles and other epidermal appendages. The dermis is a middle supporting layer containing the epidermal appendages, blood vessels, nerves and nerve endings, which are embedded in an elastocollagenous stroma produced by fibroblasts. The subcutis is the deepest layer and varies in size and content, but is usually composed mainly of adipose tissue (Fig. 18.1). The epidermis is the protective skin layer in contact with the external environment A stratified epithelium (see p. 38), the surface of the epidermis bears closely packed flat plates of protein (keratin), which form a tough, water-repellent layer (stratum corneum). Keratin is produced by the main cell type in epidermis, the keratinocyte. Note that, contrary to traditional teaching, the stratum corneum is not cellular but is composed entirely of intracytoplasmic keratin remnants bound to the skin surface after the death of the keratinocytes that produced them (see Fig. 3.28). Each keratin plate conforms roughly to the shape of the keratinocyte shortly before its death. Although normally thin, the stratum corneum is very thick in skin exposed to constant trauma, such as that on the soles and palms. Epidermis is traditionally regarded as a stratified squamous epithelium, but in fact only the most superficial two or three living cell layers of the epidermis approach a squamous or flat configuration, most keratinocytes being cuboidal or polyhedral. The surface plates of keratin, and the flat dying keratinocytes that precede them, are known as squames, and result from the maturation of the other layers of keratinocytes comprising the epidermis. These layers are the basal layer, the prickle cell layer and the granular layer (Fig. 18.2). Nomenclature of the Epidermal Layers In this book, we use the English nomenclature for the layers of the epidermis, rather than their Latin equivalents: • Basal layer ≡ stratum basale or stratum germinativum • Prickle cell layer ≡ stratum spinosum • Granular layer ≡ stratum granulosum In addition, a further layer, the stratum lucidum, is described in the skin of the sole. This refers to a narrow, pale-staining layer of compact keratin sometimes seen between the granular layer and the more intensely staining bulk of the thick keratin layer. It has no structural or functional significance and may be an artifact of staining (see Fig. 18.20). Keratinocytes are separated from the underlying support tissues of the dermis by a basement membrane The junction between the dermis and epidermis is an important area, tethering the two layers together (Fig. 18.3) and structured to minimize the risk of dermoepidermal separation by shearing forces as follows: • Tethering fibres connect the dermis and epidermis to the intervening basement membrane • The basal cell membrane of individual basal cells, and the underlying basement membrane, are convoluted (see Fig. 18.6) • There is a system of rete ridges (i.e. downgrowths of epidermis into dermis), which varies markedly from site to site. In protected areas, where the skin is not normally subjected to shearing stress (e.g. the trunk), rete ridges are barely evident and the dermoepidermal junction appears flat, but in areas constantly exposed to shearing stress (e.g. tips of fingers, palms and soles) the rete ridge system is highly developed (see Figs 18.1a, 18.5, 18.12). The basement membrane at the dermoepidermal junction can be seen to consist of three main layers, specifically: • An electron-lucent lamina lucida on the epidermal side • An electron-dense lamina densa in the middle • An ill-defined fibroreticular lamina, which contains abundant fibronectin, on the dermal side. The basal cells are tethered to the lamina densa by hemidesmosomes from which anchoring proteins cross the lamina lucida. On the dermal aspect, fine anchoring fibrils of type VII collagen attach the lower surface of the lamina densa to collagen fibres in the papillary dermis, whereas fibrillin microfibrils attach it to upper dermal elastic fibres. In addition, the zone immediately beneath the lamina densa contains abundant fibronectin (see p. 59 and Fig. 18.3c). The basal layer is the deepest cell layer of the epidermis and is responsible for the constant production of keratinocytes The basal layer cells are cuboidal or low columnar in shape and are attached to the basement membrane, which separates it from the underlying dermis, by hemidesmosomes, and to adjacent basal cells by true desmosomes. Basal cells have round or oval nuclei (Fig. 18.4) with prominent nucleoli, and their cytoplasm is rich in ribosomes and mitochondria; tonofilaments are present in small numbers. In pigmented skin the cytoplasm also contains melanin granules and lysosomes. It is in the basal layer that cells in mitosis are seen, as well as scattered non-keratinocyte cells, melanocytes and Merkel cells (see p. 376). Basal Blisters The dermoepidermal junction is a site where fluid can accumulate in sufficient quantity to lift the epidermis away from the dermis, thereby forming a basal blister. This can result from: Excessive shearing forces are the most common cause and are usually due to constant shearing friction, such as occurs with tight-fitting shoes. Structural abnormalities may be primary or secondary, and the most important primary cause is the rare inherited skin disease epidermolysis bullosa, in which the dermoepidermal junction is intrinsically weak and unable to resist even minimal shearing trauma. There are a number of different types of epidermolysis bullosa, depending on the site of separation, which can be ascertained by electron microscopy. The keratinocytes above the basal cells form the prickle cell layers The cells are polyhedral with central round nuclei and pinkish-staining cytoplasm. Prickle cells are in contact with each other by a system of intercellular bridges, formed from small cytoplasmic projections from the cell surface terminating in desmosomal junctions (Fig. 18.6; see also Fig. 3.11). The prickle cells form a layer of variable thickness, sometimes called the ‘stratum spinosum’. The cytoplasm of the prickle cells contains many tonofilaments (see Fig. 3.28), which are particularly concentrated in the cytoplasmic projections leading into the desmosomes and are more numerous in the cell layers closest to the granular layer. The cells of the upper prickle cell layer are flatter than the polyhedral cells of the deeper layers. The narrow interstices between the prickle cells are partly occupied by the cytoplasmic projections of melanocytes and Langerhans’ cells. The granular cell layer produces the surface keratin and non-wettable substances The granular keratinocytes contain small or oval haematoxyphilic round bodies (keratohyaline granules) composed of proteinaceous material containing abundant sulfur-rich amino acids (e.g. cysteine). They also contain abundant tonofibrils and small round lamellated keratinosomes or Odland bodies. In the upper layer, the cytoplasm of granular keratinocytes is largely composed of masses of keratohyaline material and tightly packed tonofibrils, with little cytosol and few cytoplasmic organelles, and at this level, the cells are flat. Death of the nucleus and cytoplasm leaves the keratohyaline and tonofibrils, which combine to form the keratin (see Fig. 3.28) of the acellular surface layer, the stratum corneum (Fig. 18.7). Keratinosomes produce a complex hydrophobic glycophospholipid, which is released when the superficial granular keratinocytes die, and probably acts as a glue, cementing together the flakes of keratin. This substance also renders the skin surface relatively non-wettable, although prolonged exposure will wash it away, permitting the keratin to absorb water, swell and soften. This feature is obvious in the skin of the hands after prolonged immersion in water, particularly when the water is hot and contains some form of lipid-dissolving detergent. Tumours of the Epidermal Cells Tumours of the epidermal cells in skin are very common, and most are associated with prolonged exposure of the skin to ultraviolet light. They are therefore most common in sun-exposed areas of skin such as the face and the backs of the hands, and particularly in the elderly or those who have had excessive sun exposure at an early age through their lifestyle. The commonest tumour is derived from the basal cells of the epidermis and is therefore called basal cell carcinoma. These tumours are malignant and, unless treated by complete excision when they are first noticed, may continue to grow slowly and destroy skin and deep tissues locally; they never metastasize to distant sites. Slightly less common are tumours derived from the keratinocytes from the stratum spinosum (prickle cell layer). These are called squamous cell carcinomas and, like basal cell carcinomas, are locally invasive and destructive, but these tumours also spread to distant sites via lymphatics. Surface keratin is constantly lost due to normal wear and tear from surface friction, washing and scrubbing Surface keratin needs to be constantly replenished from the granular layer, which in turn is constantly repopulated by cells from the prickle layer. The prickle cells are produced by proliferation of cells in the basal layer. Turnover, from basal cell to desquamated keratin, varies from site to site, being faster (i.e. 25–30 days) in traumatized areas (e.g. soles); slow turnovers range from 40 to 50 days. The turnover period is considerably shortened in some skin diseases, particularly psoriasis. In addition to keratinocytes, the epidermis also contains melanocytes, Langerhans’ cells and Merkel cells. Melanocytes produce the protective pigment melanin Melanin is largely responsible for skin colour and minimizes tissue damage by ultraviolet radiation. Melanocytes are derived from neuroectoderm and are located in the basal layer of keratinocytes in contact with the basement membrane. They are pale staining, with large ovoid nuclei and abundant cytoplasm, from which numerous long cytoplasmic processes extend into the spaces between the keratinocytes. Melanocyte cytoplasm contains characteristic membrane-bound ovoid granules (premelanosomes and melanosomes), which have a striated electron-dense core and produce melanin (Fig. 18.8c). In the production of melanin, tyrosine is converted into an intermediate pigment, which polymerizes into melanin. Melanin binds to proteins to form the active melanoprotein complex, which appears ultrastructurally as spherical masses of homogeneous electron-dense material, and often obscures the premelanosomes. Melanoprotein complexes pass along the cytoplasmic processes of the melanocyte and are transferred into the cytoplasm of basal and prickle cell layer keratinocytes, the highest concentration being in the basal layers. Melanocyte numbers remain more or less constant but their degree of activity is genetically variable, accounting for racial and individual variation in skin colour. Tumours of Melanocytes Moles or nevi are local aggregations of melanocytes in the epidermis and dermis, producing the regular, slightly raised, brown skin lesions familiar to everyone. These are local developmental abnormalities and are not true tumours. However, malignant melanocytic tumours can occasionally develop within them, the early indications that this is about to happen being changes in the shape, size and pigmentation of a nevus, which had previously been unchanged for many years. These malignant tumours of melanocytes are called malignant melanomas and may also occur in normal skin, which has no evidence of a pre-existing nevus. If recognized and excised at an early stage, malignant melanomas are curable, but once they reach a certain size and depth of invasion into the dermis, they begin to spread away from the site of origin, initially by lymphatic channels to regional lymph nodes, and eventually by the bloodstream to many different sites, such as the liver, lungs, brain, etc. Langerhans’ cells are antigen recognition cells Langerhans’ cells are located in all layers of the epidermis but are most easily seen in the prickle cell layer. They recognize antigen and are an important component of the immune system (see pp. 128-129). Like melanocytes, Langerhans’ cells have an ovoid, pale-staining nucleus surrounded by pale-staining cytoplasm from which cytoplasmic (dendritic) processes extend between the keratinocytes. Langerhans’ cell cytoplasm contains scattered characteristic Birbeck granules, which are rod-like structures with periodic cross-striations and are most numerous near the Golgi. Sometimes one end of the rod is distended to form a spherical saccule, giving the appearance of a tennis racket (Fig. 18.9d). Although present in small numbers in healthy skin, Langerhans’ cells are increased both in number, and in the extent and complexity of their dendritic processes, in many chronic inflammatory skin disorders, particularly those with an allergic or immune aetiology, such as chronic atopic dermatitis. Merkel cells are sensory receptors in the epidermis Merkel cells are scanty and difficult to demonstrate in normal skin. They are found in the basal layer and resemble melanocytes by routine light microscopy, but electron microscopy reveals rounded membrane-bound cytoplasmic neuroendocrine-type granules. Merkel cells form synaptic junctions with peripheral nerve endings at the base of the cell and also scanty desmosomal attachments to adjacent keratinocytes (Fig. 18.10). They occur either as scattered solitary cells or as aggregates, when they are associated with a so-called hair disc, located immediately beneath the basement membrane. Such aggregates are thought to be touch receptors and are sometimes called ‘tactile corpuscles’. Dermatitis (Inflammation of Skin) The skin is exposed to many damaging agents, such as chemicals and ultraviolet irradiation, which produce a wide variety of common rashes. A common type of dermatitis, seborrhoeic dermatitis, is shown in Figure 18.11. In addition, the skin reacts to internal abnormalities, and produces rashes in response to disorders, such as viral infections (e.g. measles) and drug allergies (e.g. penicillin rash). The cause of most skin diseases is not known.
Skin and Breast
Introduction
Epidermis
 Advanced Concept
Advanced Concept
 Clinical Example
Clinical Example
 Clinical Example
Clinical Example
Non-keratinizing Epidermal Cells
 Clinical Example
Clinical Example
 Clinical Example
Clinical Example