452 | Amyotrophic Lateral Sclerosis and Other Motor Neuron Diseases |
AMYOTROPHIC LATERAL SCLEROSIS
Amyotrophic lateral sclerosis (ALS) is the most common form of progressive motor neuron disease. It is a prime example of a neurodegenerative disease and is arguably the most devastating of the neurodegenerative disorders.
PATHOLOGY
The pathologic hallmark of motor neuron degenerative disorders is death of lower motor neurons (consisting of anterior horn cells in the spinal cord and their brainstem homologues innervating bulbar muscles) and upper, or corticospinal, motor neurons (originating in layer five of the motor cortex and descending via the pyramidal tract to synapse with lower motor neurons, either directly or indirectly via interneurons) (Chap. 30). Although at its onset ALS may involve selective loss of function of only upper or lower motor neurons, it ultimately causes progressive loss of both categories of motor neurons. Indeed, in the absence of clear involvement of both motor neuron types, the diagnosis of ALS is questionable. In a subset of cases, ALS arises concurrently with frontotemporal dementia (Chap. 448); in these instances, there is degeneration of frontotemporal cortical neurons and corresponding cortical atrophy.
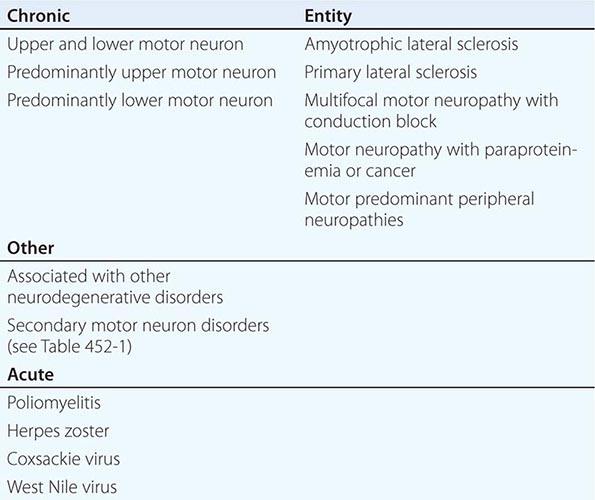
Other motor neuron diseases involve only particular subsets of motor neurons (Tables 452-1 and 452-2). Thus, in bulbar palsy and spinal muscular atrophy (SMA; also called progressive muscular atrophy), the lower motor neurons of brainstem and spinal cord, respectively, are most severely involved. By contrast, pseudobulbar palsy, primary lateral sclerosis (PLS), and familial spastic paraplegia (FSP) affect only upper motor neurons innervating the brainstem and spinal cord.
ETIOLOGY OF MOTOR NEURON DISORDERS |
SPORADIC MOTOR NEURON DISEASES |
In each of these diseases, the affected motor neurons undergo shrinkage, often with accumulation of the pigmented lipid (lipofuscin) that normally develops in these cells with advancing age. In ALS, the motor neuron cytoskeleton is typically affected early in the illness. Focal enlargements are frequent in proximal motor axons; ultrastructurally, these “spheroids” are composed of accumulations of neurofilaments and other proteins. Commonly in both sporadic and familial ALS, the affected neurons demonstrate ubiquitin-positive aggregates, typically associated with the protein TDP43 (see below). Also seen is proliferation of astroglia and microglia, the inevitable accompaniment of all degenerative processes in the central nervous system (CNS).
The death of the peripheral motor neurons in the brainstem and spinal cord leads to denervation and consequent atrophy of the corresponding muscle fibers. Histochemical and electrophysiologic evidence indicates that in the early phases of the illness denervated muscle can be reinnervated by sprouting of nearby distal motor nerve terminals, although reinnervation in this disease is considerably less extensive than in most other disorders affecting motor neurons (e.g., poliomyelitis, peripheral neuropathy). As denervation progresses, muscle atrophy is readily recognized in muscle biopsies and on clinical examination. This is the basis for the term amyotrophy. The loss of cortical motor neurons results in thinning of the corticospinal tracts that travel via the internal capsule (Fig. 452-1) and brainstem to the lateral and anterior white matter columns of the spinal cord. The loss of fibers in the lateral columns and resulting fibrillary gliosis impart a particular firmness (lateral sclerosis). A remarkable feature of the disease is the selectivity of neuronal cell death. By light microscopy, the entire sensory apparatus, the regulatory mechanisms for the control and coordination of movement, remains intact. Except in cases of frontotemporal dementia, the components of the brain required for cognitive processing are also preserved. However, immunostaining indicates that neurons bearing ubiquitin, a marker for degeneration, are also detected in nonmotor systems. Moreover, studies of glucose metabolism in the illness also indicate that there is neuronal dysfunction outside of the motor system. Within the motor system, there is some selectivity of involvement. Thus, motor neurons required for ocular motility remain unaffected, as do the parasympathetic neurons in the sacral spinal cord (the nucleus of Onufrowicz, or Onuf) that innervate the sphincters of the bowel and bladder.
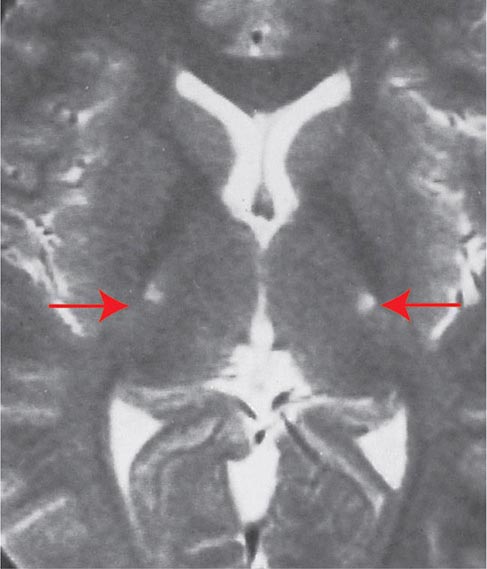
FIGURE 452-1 Amyotrophic lateral sclerosis. Axial T2-weighted magnetic resonance imaging (MRI) scan through the lateral ventricles of the brain reveals abnormal high signal intensity within the corticospinal tracts (arrows). This MRI feature represents an increase in water content in myelin tracts undergoing Wallerian degeneration secondary to cortical motor neuronal loss. This finding is commonly present in ALS, but can also be seen in AIDS-related encephalopathy, infarction, or other disease processes that produce corticospinal neuronal loss in a symmetric fashion.
CLINICAL MANIFESTATIONS
The manifestations of ALS are somewhat variable depending on whether corticospinal neurons or lower motor neurons in the brainstem and spinal cord are more prominently involved. With lower motor neuron dysfunction and early denervation, typically the first evidence of the disease is insidiously developing asymmetric weakness, usually first evident distally in one of the limbs. A detailed history often discloses recent development of cramping with volitional movements, typically in the early hours of the morning (e.g., while stretching in bed). Weakness caused by denervation is associated with progressive wasting and atrophy of muscles and, particularly early in the illness, spontaneous twitching of motor units, or fasciculations. In the hands, a preponderance of extensor over flexor weakness is common. When the initial denervation involves bulbar rather than limb muscles, the problem at onset is difficulty with chewing, swallowing, and movements of the face and tongue. Early involvement of the muscles of respiration may lead to death before the disease is far advanced elsewhere. With prominent corticospinal involvement, there is hyperactivity of the muscle-stretch reflexes (tendon jerks) and, often, spastic resistance to passive movements of the affected limbs. Patients with significant reflex hyperactivity complain of muscle stiffness often out of proportion to weakness. Degeneration of the corticobulbar projections innervating the brainstem results in dysarthria and exaggeration of the motor expressions of emotion. The latter leads to involuntary excess in weeping or laughing (pseudobulbar affect).
Virtually any muscle group may be the first to show signs of disease, but, as time passes, more and more muscles become involved until ultimately the disorder takes on a symmetric distribution in all regions. It is characteristic of ALS that, regardless of whether the initial disease involves upper or lower motor neurons, both will eventually be implicated. Even in the late stages of the illness, sensory, bowel and bladder, and cognitive functions are preserved. Even when there is severe brainstem disease, ocular motility is spared until the very late stages of the illness. As noted, in some cases (particularly those that are familial), ALS develops concurrently with frontotemporal dementia, characterized by early behavioral abnormalities with prominent behavioral features indicative of frontal lobe dysfunction.
A committee of the World Federation of Neurology has established diagnostic guidelines for ALS. Essential for the diagnosis is simultaneous upper and lower motor neuron involvement with progressive weakness and the exclusion of all alternative diagnoses. The disorder is ranked as “definite” ALS when three or four of the following are involved: bulbar, cervical, thoracic, and lumbosacral motor neurons. When two sites are involved, the diagnosis is “probable,” and when only one site is implicated, the diagnosis is “possible.” An exception is made for those who have progressive upper and lower motor neuron signs at only one site and a mutation in the gene encoding superoxide dismutase (SOD1; see below).
EPIDEMIOLOGY
The illness is relentlessly progressive, leading to death from respiratory paralysis; the median survival is from 3–5 years. There are very rare reports of stabilization or even regression of ALS. In most societies, there is an incidence of 1–3 per 100,000 and a prevalence of 3–5 per 100,000. It is striking that about 1 in 1000 adult deaths in North America and Western Europe (and probably elsewhere) are due to ALS; this finding predicts that some 300,000 individuals now alive in the United States will die of ALS. Several endemic foci of higher prevalence exist in the western Pacific (e.g., in specific regions of Guam or Papua New Guinea). In the United States and Europe, males are somewhat more frequently affected than females. Epidemiologic studies have incriminated risk factors for this disease including exposure to pesticides and insecticides, smoking, and, in one report, service in the military. Although ALS is overwhelmingly a sporadic disorder, some 5–10% of cases are inherited as an autosomal dominant trait.
FAMILIAL ALS
Several forms of selective motor neuron disease are inheritable (Table 452-3). Familial ALS (FALS) involves both corticospinal and lower motor neurons. Apart from its inheritance as an autosomal dominant trait, it is clinically indistinguishable from sporadic ALS. Genetic studies have identified mutations in multiple genes, including those encoding the protein C9orf 72 (open reading frame 72 on chromosome 9), cytosolic enzyme SOD1 (superoxide dismutase), the RNA binding proteins TDP43 (encoded by the TAR DNA binding protein gene), and FUS/TLS (fused in sarcoma/translocated in liposarcoma), as the most common causes of FALS. Mutations in C9orf72 account for ~45–50% of FALS and perhaps 4–5% of sporadic ALS cases. Mutations in SOD1 explain another 20% of cases of FALS, whereas TDP43 and FUS/TLS each represent about 5% of familial cases. It has recently been reported that ~1–2% of cases are caused by mutations in genes encoding the proteins optineuron and profilin-1 as well.
GENETIC MOTOR NEURON DISEASES |
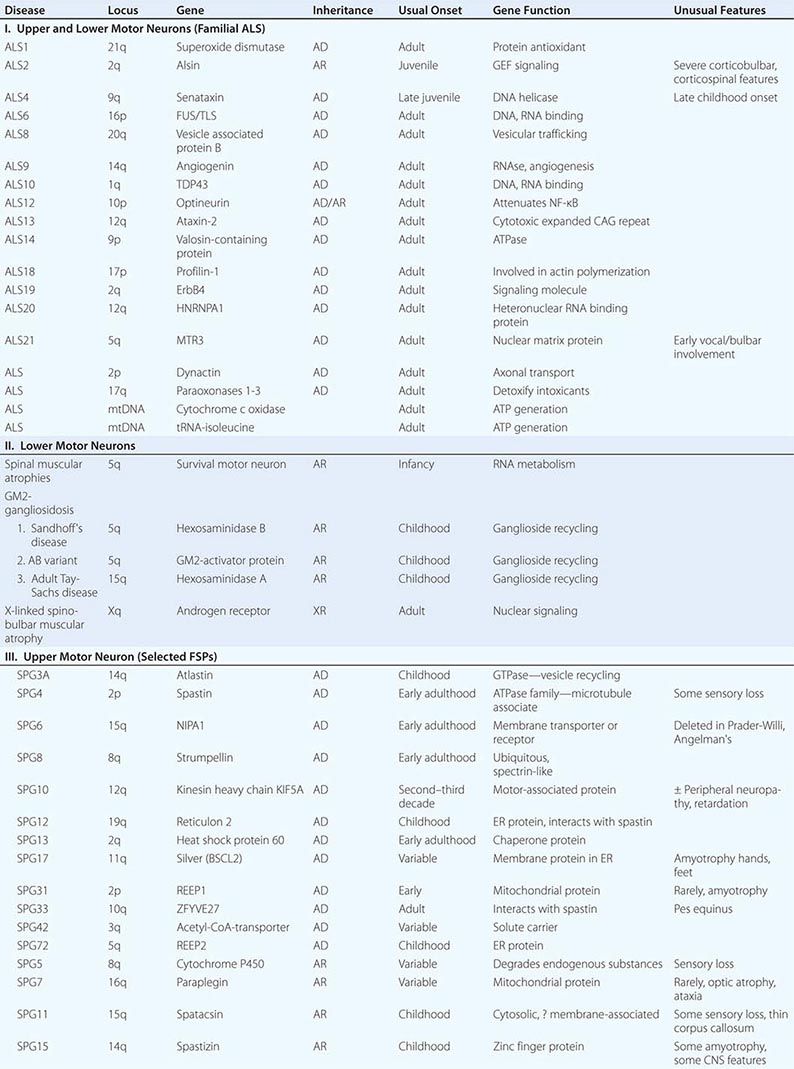
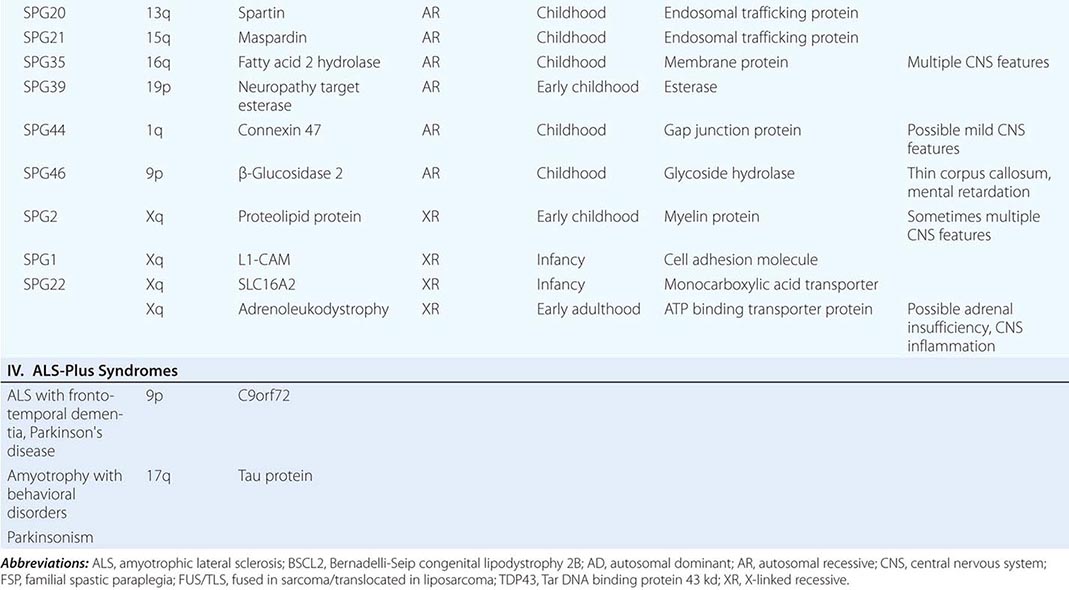
Rare mutations in other genes are also clearly implicated in ALS-like diseases. Thus, a familial, dominantly inherited motor disorder that in some individuals closely mimics the ALS phenotype arises from mutations in a gene that encodes a vesicle-binding protein. A predominantly lower motor neuron disease with early hoarseness due to laryngeal dysfunction has been ascribed to mutations in the gene encoding the cellular accessory motor protein dynactin. Mutations in senataxin, a helicase, cause an early adult-onset, slowly evolving ALS variant. Kennedy’s syndrome is an X-linked, adult-onset disorder that may mimic ALS, as described below.
Genetic analyses are also beginning to illuminate the pathogenesis of some childhood-onset motor neuron diseases. For example, a slowly disabling degenerative, predominantly upper motor neuron disease that starts in the first decade is caused by mutations in a gene that expresses a novel signaling molecule with properties of a guanine-exchange factor, termed alsin.
DIFFERENTIAL DIAGNOSIS
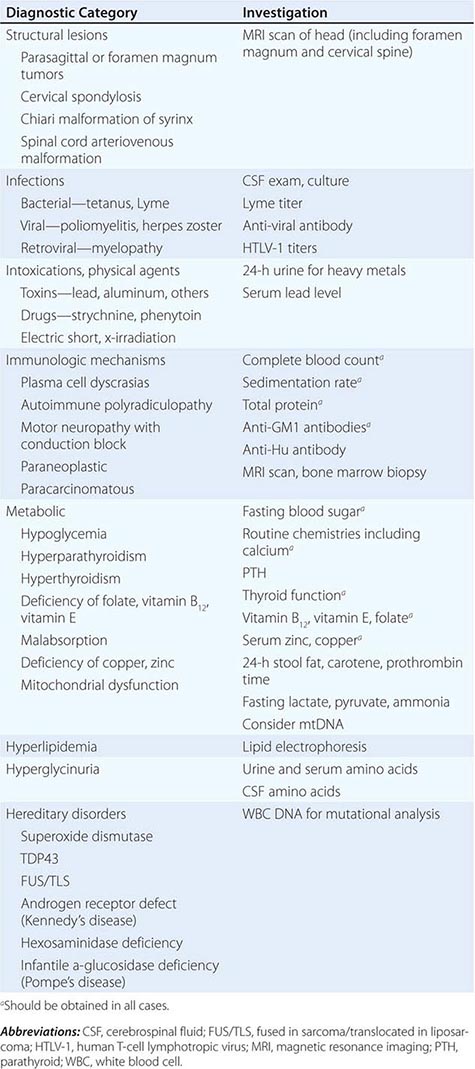
Because ALS is currently untreatable, it is imperative that potentially remediable causes of motor neuron dysfunction be excluded (Table 452-1). This is particularly true in cases that are atypical by virtue of (1) restriction to either upper or lower motor neurons, (2) involvement of neurons other than motor neurons, and (3) evidence of motor neuronal conduction block on electrophysiologic testing. Compression of the cervical spinal cord or cervicomedullary junction from tumors in the cervical regions or at the foramen magnum or from cervical spondylosis with osteophytes projecting into the vertebral canal can produce weakness, wasting, and fasciculations in the upper limbs and spasticity in the legs, closely resembling ALS. The absence of cranial nerve involvement may be helpful in differentiation, although some foramen magnum lesions may compress the twelfth cranial (hypoglossal) nerve, with resulting paralysis of the tongue. Absence of pain or of sensory changes, normal bowel and bladder function, normal roentgenographic studies of the spine, and normal cerebrospinal fluid (CSF) all favor ALS. Where doubt exists, magnetic resonance imaging (MRI) scans and contrast myelography should be performed to visualize the cervical spinal cord.
Another important entity in the differential diagnosis of ALS is multifocal motor neuropathy with conduction block (MMCB), discussed below. A diffuse, lower motor axonal neuropathy mimicking ALS sometimes evolves in association with hematopoietic disorders such as lymphoma or multiple myeloma. In this clinical setting, the presence of an M-component in serum should prompt consideration of a bone marrow biopsy. Lyme disease (Chap. 210) may also cause an axonal, lower motor neuropathy, although typically with intense proximal limb pain and a CSF pleocytosis.
Other treatable disorders that occasionally mimic ALS are chronic lead poisoning and thyrotoxicosis. These disorders may be suggested by the patient’s social or occupational history or by unusual clinical features. When the family history is positive, disorders involving the genes encoding C9orf72, cytosolic SOD1, TDP43, FUS/TLS, and adult hexosaminidase A or α-glucosidase deficiency must be excluded (Chap. 432e). These are readily identified by appropriate laboratory tests. Benign fasciculations are occasionally a source of concern because on inspection they resemble the fascicular twitchings that accompany motor neuron degeneration. The absence of weakness, atrophy, or denervation phenomena on electrophysiologic examination usually excludes ALS or other serious neurologic disease. Patients who have recovered from poliomyelitis may experience a delayed deterioration of motor neurons that presents clinically with progressive weakness, atrophy, and fasciculations. Its cause is unknown, but it is thought to reflect sublethal prior injury to motor neurons by poliovirus (Chap. 228).
Rarely, ALS develops concurrently with features indicative of more widespread neurodegeneration. Thus, one infrequently encounters otherwise typical ALS patients with a parkinsonian movement disorder or frontotemporal dementia, particularly in instances of C9orf72 mutations, which strongly suggests that the simultaneous occurrence of two disorders is a direct consequence of the gene mutation. As another example, prominent amyotrophy has been described as a dominantly inherited disorder in individuals with bizarre behavior and a movement disorder suggestive of parkinsonism; many such cases have now been ascribed to mutations that alter the expression of tau protein in brain (Chap. 448). In other cases, ALS develops simultaneously with a striking frontotemporal dementia. An ALS-like disorder has also been described in some individuals with chronic traumatic encephalopathy, associated with deposition of TDP43 and neurofibrillary tangles in motor neurons.
PATHOGENESIS
The cause of sporadic ALS is not well defined. Several mechanisms that impair motor neuron viability have been elucidated in mice and rats induced to develop motor neuron disease by SOD1 transgenes with ALS-associated mutations. One may loosely group the genetic causes of ALS into three categories. In one group, the primarily problem is inherent instability of the mutant proteins, with subsequent perturbations in protein degradation (SOD1, ubiquilin-1 and -2, p62). In the second, most rapidly growing category, the causative mutant genes perturb RNA processing, transport, and metabolism (C9orf73, TDP43, FUS). In the case of C9orf72, the molecular pathology is an expansion of an intronic hexanucleotide repeat (-GGGGCC-) beyond an upper normal of 30 repeats to hundreds or more repeats. As observed in other intronic repeat disorders such as myotonic dystrophy (Chap. 462e) and spinocerebellar atrophy type 8 (Chap. 450), data suggest that the expanded intronic repeats generate expanded RNA repeats that form intranuclear foci and confer toxicity by sequestering transcription factors or by undergoing noncanonical protein translation across all possible reading frames of the expanded RNA tracts. TDP43 and FUS are multifunctional proteins that bind RNA and DNA and shuttle between the nucleus and the cytoplasm, playing multiple roles in the control of cell proliferation, DNA repair and transcription, and gene translation, both in the cytoplasm and locally in dendritic spines in response to electrical activity. How mutations in FUS/TLS provoke motor neuron cell death is not clear, although this may represent loss of function of FUS/TLS in the nucleus or an acquired, toxic function of the mutant proteins in the cytosol. In the third group of ALS genes, the primary problem is defective axonal cytoskeleton and transport (dynactin, profilin-1). It is striking that variants in other genes (e.g., EphA4) influence survival in ALS but not ALS susceptibility. Beyond the upstream, primary defects, it is also evident that the ultimate neuronal cell death process is complex involving multiple cellular processes that accelerate cell death. These include but are not limited to excitotoxicity, impairment of axonal transport, oxidative stress, activation of endoplasmic reticulum stress and the unfolded protein response, and mitochondrial dysfunction.
Multiple recent studies have convincingly demonstrated that nonneuronal cells importantly influence the disease course, at least in ALS transgenic mice. A striking additional finding in neurodegenerative disorders is that miscreant proteins arising from gene defects in familial forms of these diseases are often implicated in sporadic forms of the same disorder. For example, germline mutations in the genes encoding β-amyloid and α-synuclein cause familial forms of Alzheimer’s and Parkinson’s diseases, and posttranslational, noninherited abnormalities in these proteins are also central to sporadic Alzheimer’s and Parkinson’s diseases. Analogously, recent reports propose that nonheritable, posttranslational modifications in SOD1 are pathogenic in sporadic ALS.
OTHER MOTOR NEURON DISEASES
SELECTED LOWER MOTOR NEURON DISORDERS
In these motor neuron diseases, the peripheral motor neurons are affected without evidence of involvement of the corticospinal motor system (Tables 452-1, 452-2, and 452-3).
X-Linked Spinobulbar Muscular Atrophy (Kennedy’s Disease) This is an X-linked lower motor neuron disorder in which progressive weakness and wasting of limb and bulbar muscles begins in males in mid-adult life and is conjoined with androgen insensitivity manifested by gynecomastia and reduced fertility (Chap. 411). In addition to gynecomastia, which may be subtle, two findings distinguishing this disorder from ALS are the absence of signs of pyramidal tract disease (spasticity) and the presence of a subtle sensory neuropathy in some patients. The underlying molecular defect is an expanded trinucleotide repeat (-CAG-) in the first exon of the androgen receptor gene on the × chromosome. DNA testing is available. An inverse correlation appears to exist between the number of -CAG- repeats and the age of onset of the disease.
Adult Tay-Sachs Disease Several reports have described adult-onset, predominantly lower motor neuropathies arising from deficiency of the enzymeβ-hexosaminidase (hex A). These tend to be distinguishable from ALS because they are very slowly progressive; dysarthria and radiographically evident cerebellar atrophy may be prominent. In rare cases, spasticity may also be present, although it is generally absent (Chap. 432e).
Spinal Muscular Atrophy The SMAs are a family of selective lower motor neuron diseases of early onset. Despite some phenotypic variability (largely in age of onset), the defect in the majority of families with SMA maps to a locus on chromosome 5 encoding a putative motor neuron survival protein (SMN, for survival motor neuron) that is important in the formation and trafficking of RNA complexes across the nuclear membrane. Neuropathologically these disorders are characterized by extensive loss of large motor neurons; muscle biopsy reveals evidence of denervation atrophy. Several clinical forms exist.
Infantile SMA (SMA I, Werdnig-Hoffmann disease) has the earliest onset and most rapidly fatal course. In some instances it is apparent even before birth, as indicated by decreased fetal movements late in the third trimester. Though alert, afflicted infants are weak and floppy (hypotonic) and lack muscle stretch reflexes. Death generally ensues within the first year of life. Chronic childhood SMA (SMA II) begins later in childhood and evolves with a more slowly progressive course. Juvenile SMA (SMA III, Kugelberg-Welander disease) manifests during late childhood and runs a slow, indolent course. Unlike most denervating diseases, in this chronic disorder, weakness is greatest in the proximal muscles; indeed, the pattern of clinical weakness can suggest a primary myopathy such as limb-girdle dystrophy. Electrophysiologic and muscle biopsy evidence of denervation distinguish SMA III from the myopathic syndromes. There is no primary therapy for SMA, although remarkable recent experimental data indicate that it may be possible to deliver the missing SMN gene to motor neurons using intravenously or intrathecally delivered adeno-associated viruses (e.g., AAV9) immediately after birth.
Multifocal Motor Neuropathy with Conduction Block In this disorder lower motor neuron function is regionally and chronically disrupted by remarkably focal blocks in conduction. Many cases have elevated serum titers of mono- and polyclonal antibodies to ganglioside GM1; it is hypothesized that the antibodies produce selective, focal, paranodal demyelination of motor neurons. MMCB is not typically associated with corticospinal signs. In contrast with ALS, MMCB may respond dramatically to therapy such as IV immunoglobulin or chemotherapy; thus, it is imperative that MMCB be excluded when considering a diagnosis of ALS.
Other Forms of Lower Motor Neuron Disease In individual families, other syndromes characterized by selective lower motor neuron dysfunction in an SMA-like pattern have been described. There are rare X-linked and autosomal dominant forms of apparent SMA. There is an ALS variant of juvenile onset, the Fazio-Londe syndrome, that involves mainly the musculature innervated by the brainstem. A component of lower motor neuron dysfunction is also found in degenerative disorders such as Machado-Joseph disease and the related olivopontocerebellar degenerations (Chap. 450).
SELECTED DISORDERS OF THE UPPER MOTOR NEURON
Primary Lateral Sclerosis This exceedingly rare disorder arises sporadically in adults in mid to late life. Clinically PLS is characterized by progressive spastic weakness of the limbs, preceded or followed by spastic dysarthria and dysphagia, indicating combined involvement of the corticospinal and corticobulbar tracts. Fasciculations, amyotrophy, and sensory changes are absent; neither electromyography nor muscle biopsy shows denervation. On neuropathologic examination, there is selective loss of the large pyramidal cells in the precentral gyrus and degeneration of the corticospinal and corticobulbar projections. The peripheral motor neurons and other neuronal systems are spared. The course of PLS is variable; although long-term survival is documented, the course may be as aggressive as in ALS, with ~3-year survival from onset to death. Early in its course, PLS raises the question of multiple sclerosis or other demyelinating diseases such as adrenoleukodystrophy as diagnostic considerations (Chap. 458). A myelopathy suggestive of PLS is infrequently seen with infection with the retrovirus human T cell lymphotropic virus 1 (HTLV-1) (Chap. 456). The clinical course and laboratory testing will distinguish these possibilities.
Familial Spastic Paraplegia In its pure form, FSP is usually transmitted as an autosomal trait; most adult-onset cases are dominantly inherited. Symptoms usually begin in the third or fourth decade, presenting as progressive spastic weakness beginning in the distal lower extremities; however, there are variants with onset so early that the differential diagnosis includes cerebral palsy. FSP typically has a long survival, presumably because respiratory function is spared. Late in the illness, there may be urinary urgency and incontinence and sometimes fecal incontinence; sexual function tends to be preserved.
In pure forms of FSP, the spastic leg weakness is often accompanied by posterior column (vibration and position) abnormalities and disturbance of bowel and bladder function. Some family members may have spasticity without clinical symptoms.
By contrast, particularly when recessively inherited, FSP may have complex or complicated forms in which altered corticospinal and dorsal column function is accompanied by significant involvement of other regions of the nervous system, including amyotrophy, mental retardation, optic atrophy, and sensory neuropathy.
Neuropathologically, in FSP, there is degeneration of the corticospinal tracts, which appear nearly normal in the brainstem but show increasing atrophy at more caudal levels in the spinal cord; in effect, the pathologic picture is of a dying-back or distal axonopathy of long neuronal fibers within the CNS.
Defects at numerous loci underlie both dominantly and recessively inherited forms of FSP (Table 452-3). More than 30 FSP genes have now been identified. The gene most commonly implicated in dominantly inherited FSP is spastin, which encodes a microtubule interacting protein. The most common childhood-onset dominant form arises from mutations in the atlastin gene. A kinesin heavy-chain protein implicated in microtubule motor function was found to be defective in a family with dominantly inherited FSP of variable-onset age.
An infantile-onset form of X-linked, recessive FSP arises from mutations in the gene for myelin proteolipid protein. This is an example of rather striking allelic variation, as most other mutations in the same gene cause not FSP but Pelizaeus-Merzbacher disease, a widespread disorder of CNS myelin. Another recessive variant is caused by defects in the paraplegin gene. Paraplegin has homology to metalloproteases that are important in mitochondrial function in yeast.
WEBSITES
Several websites provide valuable information on ALS including those offered by the Muscular Dystrophy Association (www.mdausa.org), the Amyotrophic Lateral Sclerosis Association (www.alsa.org), and the World Federation of Neurology and the Neuromuscular Unit at Washington University in St. Louis (www.neuro.wustl.edu).
453e | Prion Diseases |
Prions are proteins that adopt an alternative conformation, which becomes self-propagating. Some prions cause degeneration of the central nervous system (CNS). Once relegated to causing a group of rare disorders of the CNS such as Creutzfeldt-Jakob disease (CJD), prions—as mounting evidence shows—also appear to play a key role in more common illnesses such as Alzheimer’s disease (AD) and Parkinson’s disease (PD). While CJD is caused by the accumulation of PrPSc, increasing data argue that Aβ prions cause AD, α-synuclein prions cause PD, and tau prions cause the frontotemporal dementias (FTDs). In this chapter, we confine our discussion to CJD, which typically presents with a rapidly progressive dementia as well as motor abnormalities. The illness is relentlessly progressive and generally causes death within 9 months of onset. Most CJD patients are between 50 and 75 years of age; however, patients as young as 17 and as old as 83 have been recorded.
CJD is one malady in a group of disorders caused by prions composed of the prion protein (PrP). PrP prions reproduce by binding to the normal, cellular isoform of the prion protein (PrPC) and stimulating conversion of PrPC into the disease-causing isoform PrPSc. PrPC is rich in α-helix and has little β-structure, whereas PrPSc has less α-helix and a high amount of β-structure (Fig. 453e-1). This α-to-β structural transition in PrP is the fundamental event underlying this group of prion diseases (Table 453e-1).
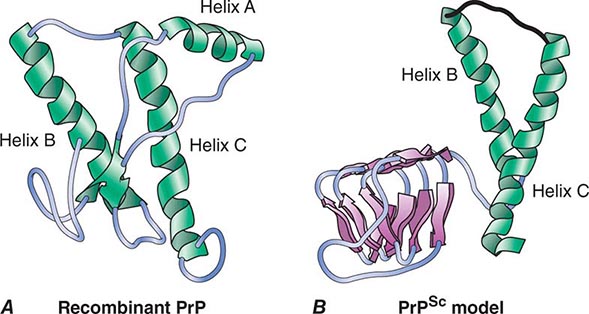
FIGURE 453e-1 Structures of prion proteins. A. NMR structure of Syrian hamster recombinant (rec) PrP(90–231). Presumably, the structure of the α-helical form of recPrP(90–231) resembles that of PrPC. recPrP(90–231) is viewed from the interface where PrPSc is thought to bind to PrPC. Shown are: α-helices A (residues 144–157), B (172–193), and C (200–227). Flat ribbons depict β-strands S1 (129–131) and S2 (161–163). B. Structural model of PrPSc. The 90–160 region has been modeled onto a β-helical architecture while the COOH terminal helices B and C are preserved as in PrPC.
GLOSSARY OF PRION TERMINOLOGY |

Four new concepts have emerged from studies of prions: (1) Prions are the only known transmissible pathogens that are devoid of nucleic acid; all other infectious agents possess genomes composed of either RNA or DNA that direct the synthesis of their progeny. (2) Prion diseases may be manifest as infectious, genetic, and sporadic disorders; no other group of illnesses with a single etiology presents with such a wide spectrum of clinical manifestations. (3) Prion diseases result from the accumulation of PrPSc, the conformation of which differs substantially from that of its precursor, PrPC. (4) Distinct strains of prions exhibit different biologic properties, which are epigenetically inherited. In other words, PrPSc can exist in a variety of different conformations, many of which seem to specify particular disease phenotypes.
How a specific conformation of a PrPSc molecule is imparted to PrPC during prion replication to produce nascent PrPSc with the same conformation is unknown. Additionally, it is unclear what factors determine where in the CNS a particular PrPSc molecule will be deposited.
SPECTRUM OF PRION DISEASES
The sporadic form of CJD is the most common prion disorder in humans. Sporadic CJD (sCJD) accounts for ~85% of all cases of human PrP prion disease, whereas inherited prion diseases account for 10–15% of all cases (Table 453e-2). Familial CJD (fCJD), Gerstmann-Sträussler-Scheinker (GSS) disease, and fatal familial insomnia (FFI) are all dominantly inherited prion diseases that are caused by mutations in the PrP gene.
THE PrP PRION DISEASES |
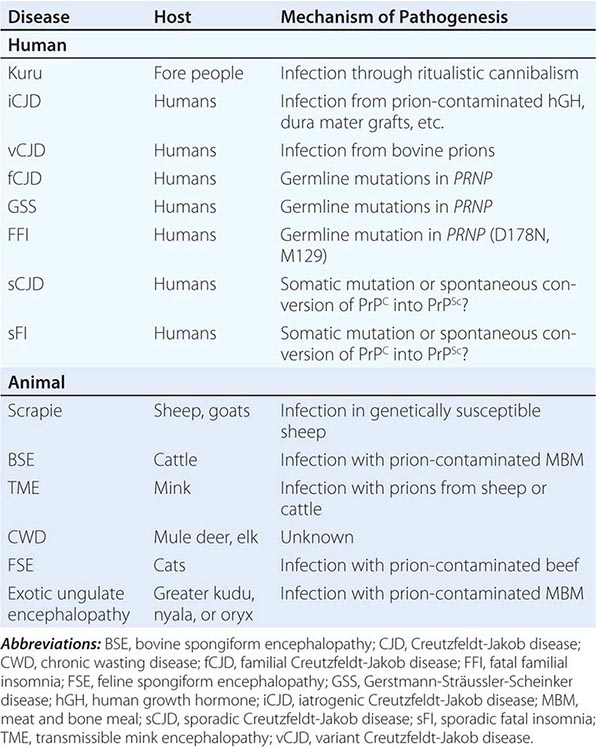
 Although infectious PrP prion diseases account for <1% of all cases and infection does not seem to play an important role in the natural history of these illnesses, the transmissibility of prions is an important biologic feature. Kuru of the Fore people of New Guinea is thought to have resulted from the consumption of brains from dead relatives during ritualistic cannibalism. With the cessation of ritualistic cannibalism in the late 1950s, kuru has nearly disappeared, with the exception of a few recent patients exhibiting incubation periods of >40 years. Iatrogenic CJD (iCJD) seems to be the result of the accidental inoculation of patients with prions. Variant CJD (vCJD) in teenagers and young adults in Europe is the result of exposure to tainted beef from cattle with bovine spongiform encephalopathy (BSE). Although occasional cases of iatrogenic CJD still occur, this form of CJD is currently on the decline due to public health measures aimed at preventing the spread of PrP prions.
Although infectious PrP prion diseases account for <1% of all cases and infection does not seem to play an important role in the natural history of these illnesses, the transmissibility of prions is an important biologic feature. Kuru of the Fore people of New Guinea is thought to have resulted from the consumption of brains from dead relatives during ritualistic cannibalism. With the cessation of ritualistic cannibalism in the late 1950s, kuru has nearly disappeared, with the exception of a few recent patients exhibiting incubation periods of >40 years. Iatrogenic CJD (iCJD) seems to be the result of the accidental inoculation of patients with prions. Variant CJD (vCJD) in teenagers and young adults in Europe is the result of exposure to tainted beef from cattle with bovine spongiform encephalopathy (BSE). Although occasional cases of iatrogenic CJD still occur, this form of CJD is currently on the decline due to public health measures aimed at preventing the spread of PrP prions.
Six diseases of animals are caused by prions (Table 453e-2). Scrapie of sheep and goats is the prototypic prion disease. Mink encephalopathy, BSE, feline spongiform encephalopathy, and exotic ungulate encephalopathy are all thought to occur after the consumption of prion-infected foodstuffs. The BSE epidemic emerged in Britain in the late 1980s and was shown to be due to industrial cannibalism. Whether BSE began as a sporadic case of BSE in a cow or started with scrapie in sheep is unknown. The origin of chronic wasting disease (CWD), a prion disease endemic in deer and elk in regions of North America, is uncertain. In contrast to other prion diseases, CWD is highly communicable. Feces from asymptomatic, infected cervids contain prions that are likely to be responsible for the spread of CWD.
EPIDEMIOLOGY
CJD is found throughout the world. The incidence of sCJD is approximately one case per million population, and thus it accounts for approximately 1 in every 10,000 deaths. Because sCJD is an age-dependent neurodegenerative disease, its incidence is expected to increase steadily as older segments of populations in developed and developing countries continue to expand. Although many geographic clusters of CJD have been reported, each has been shown to segregate with a PrP gene mutation. Attempts to identify common exposure to some etiologic agent have been unsuccessful for both the sporadic and familial cases. Ingestion of scrapie-infected sheep or goat meat as a cause of CJD in humans has not been demonstrated by epidemiologic studies, although speculation about this potential route of inoculation continues. Of particular interest are deer hunters who develop CJD, because up to 90% of culled deer in some game herds have been shown to harbor CWD prions. Whether prion disease in deer or elk has passed to cows, sheep, or directly to humans remains unknown. Studies with rodents demonstrate that oral infection with prions can occur, but the process is inefficient compared to intracerebral inoculation.
PATHOGENESIS
The human prion diseases were initially classified as neurodegenerative disorders of unknown etiology on the basis of pathologic changes being confined to the CNS. With the transmission of kuru and CJD to apes, investigators began to view these diseases as infectious CNS illnesses caused by slow viruses. Even though the familial nature of a subset of CJD cases was well described, the significance of this observation became more obscure with the transmission of CJD to animals. Eventually the meaning of heritable CJD became clear with the discovery of mutations in the PRNP gene of these patients. The prion concept explains how a disease can manifest as a heritable as well as an infectious illness. Moreover, the hallmark of all prion diseases, whether sporadic, dominantly inherited, or acquired by infection, is that they involve the aberrant metabolism of PrP.
A major feature that distinguishes prions from viruses is the finding that both PrP isoforms are encoded by a chromosomal gene. In humans, the PrP gene is designated PRNP and is located on the short arm of chromosome 20. Limited proteolysis of PrPSc produces a smaller, protease-resistant molecule of ~142 amino acids designated PrP 27-30; PrPC is completely hydrolyzed under the same conditions (Fig. 453e-2). In the presence of detergent, PrP 27-30 polymerizes into amyloid. Prion rods formed by limited proteolysis and detergent extraction are indistinguishable from the filaments that aggregate to form PrP amyloid plaques in the CNS. Both the rods and the PrP amyloid filaments found in brain tissue exhibit similar ultrastructural morphology and green-gold birefringence after staining with Congo red dye.
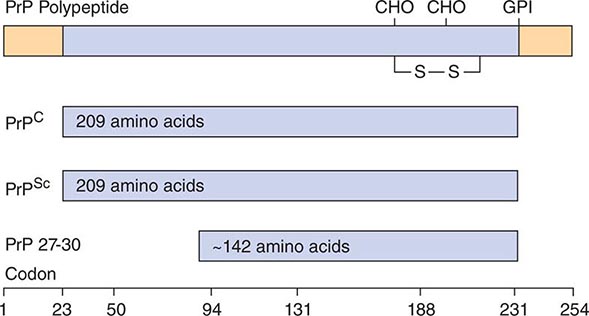
FIGURE 453e-2 Prion protein isoforms. Bar diagram of Syrian hamster PrP, which consists of 254 amino acids. After processing of the NH2 and COOH termini, both PrPC and PrPSc consist of 209 residues. After limited proteolysis, the NH2 terminus of PrPSc is truncated to form PrP 27–30 composed of ~142 amino acids. GPI, glycosylphosphatidyl inositol anchor attachment site; S—S, disulfide bond; CHO, N-linked sugars.
Prion Strains Distinct strains of prions exhibit different biologic properties, which are epigenetically heritable. The existence of prion strains raised the question of how heritable biologic information can be enciphered in a molecule other than nucleic acid. Various strains of prions have been defined by incubation times and the distribution of neuronal vacuolation. Subsequently, the patterns of PrPSc deposition were found to correlate with vacuolation profiles, and these patterns were also used to characterize prion strains.
Persuasive evidence that strain-specific information is enciphered in the tertiary structure of PrPSc comes from transmission of two different inherited human prion diseases to mice expressing a chimeric human-mouse PrP transgene. In FFI, the protease-resistant fragment of PrPSc after deglycosylation has a molecular mass of 19 kDa, whereas in fCJD and most sporadic prion diseases, it is 21 kDa (Table 453e-3). This difference in molecular mass was shown to be due to different sites of proteolytic cleavage at the NH2 termini of the two human PrPSc molecules, reflecting different tertiary structures. These distinct conformations were not unexpected because the amino acid sequences of the PrPs differ.
DISTINCT PRION STRAINS GENERATED IN HUMANS WITH INHERITED PRION DISEASES AND TRANSMITTED TO TRANSGENIC MICEa |
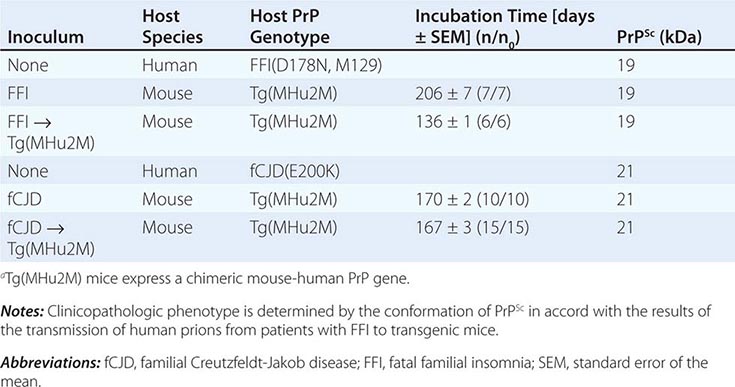
Extracts from the brains of patients with FFI transmitted disease into mice expressing a chimeric human-mouse PrP transgene and induced formation of the 19-kDa PrPSc, whereas brain extracts from FCJD and sCJD patients produced the 21-kDa PrPSc in mice expressing the same transgene. On second passage, these differences were maintained, demonstrating that chimeric PrPSc can exist in two different conformations based on the sizes of the protease-resistant fragments, even though the amino acid sequence of PrPSc is invariant.
This analysis was extended when patients with sporadic fatal insomnia (sFI) were identified. Although they did not carry a PRNP gene mutation, the patients demonstrated a clinical and pathologic phenotype that was indistinguishable from that of patients with FFI. Furthermore, 19-kDa PrPSc was found in their brains, and on passage of prion disease to mice expressing a chimeric human-mouse PrP transgene, 19-kDa PrPSc was also found. These findings indicate that the disease phenotype is dictated by the conformation of PrPSc and not the amino acid sequence. PrPSc acts as a template for the conversion of PrPC into nascent PrPSc. On the passage of prions into mice expressing a chimeric hamster-mouse PrP transgene, a change in the conformation of PrPSc was accompanied by the emergence of a new strain of prions.
Many new strains of prions were generated using recombinant (rec) PrP produced in bacteria; recPrP was polymerized into amyloid fibrils and inoculated into transgenic mice expressing high levels of wild-type mouse PrPC; approximately 500 days later, the mice died of prion disease. The incubation times of the “synthetic prions” in mice were dependent on the conditions used for polymerization of the amyloid fibrils. Highly stable amyloids gave rise to stable prions with long incubation times; low-stability amyloids led to prions with short incubation times. Amyloids of intermediate stability gave rise to prions with intermediate stabilities and intermediate incubation times. Such findings are consistent with earlier studies showing that the incubation times of synthetic and naturally occurring prions are directly proportional to the stability of the prion.
Species Barrier Studies on the role of the primary and tertiary structures of PrP in the transmission of prion disease have given new insights into the pathogenesis of these maladies. The amino acid sequence of PrP encodes the species of the prion, and the prion derives its PrPSc sequence from the last mammal in which it was passaged. While the primary structure of PrP is likely to be the most important or even sole determinant of the tertiary structure of PrPC, PrPSc seems to function as a template in determining the tertiary structure of nascent PrPSc molecules as they are formed from PrPC. In turn, prion diversity appears to be enciphered in the conformation of PrPSc, and thus prion strains seem to represent different conformers of PrPSc.
In general, transmission of PrP prion disease from one species to another is inefficient, in that not all intracerebrally inoculated animals develop disease, and those that fall ill do so only after long incubation times that can approach the natural life span of the animal. This “species barrier” to transmission is correlated with the degree of similarity between the amino acid sequences of PrPC in the inoculated host and of PrPSc in the prion inoculum. The importance of sequence similarity between the host and donor PrP argues that PrPC directly interacts with PrPSc in the prion conversion process.
SPORADIC AND INHERITED PrP PRION DISEASES
Several different scenarios might explain the initiation of sporadic prion disease: (1) A somatic mutation may be the cause and thus follow a path similar to that for germline mutations in inherited disease. In this situation, the mutant PrPSc must be capable of targeting wild-type PrPC, a process known to be possible for some mutations but less likely for others. (2) The activation energy barrier separating wild-type PrPC from PrPSc could be crossed on rare occasions when viewed in the context of a population. Most individuals would be spared, while presentations in the elderly with an incidence of ~1 per million would be seen. (3) PrPSc may be present at low levels in some normal cells, where it performs some important, as yet unknown, function. The level of PrPSc in such cells is hypothesized to be sufficiently low as to be not detected by routine bioassay. In some altered metabolic states, the cellular mechanisms for clearing PrPSc might become compromised, and the rate of PrPSc formation would then begin to exceed the capacity of the cell to clear it. The third possible mechanism is attractive because it suggests PrPSc is not simply a misfolded protein, as proposed for the first and second mechanisms, but that it is an alternatively folded molecule with a function. Moreover, the multitude of conformational states that PrPSc can adopt, as described above, raises the possibility that PrPSc or another prion-like protein might function in a process like short-term memory where information storage occurs in the absence of new protein synthesis.
More than 40 different mutations resulting in nonconservative substitutions in the human PRNP gene have been found to segregate with inherited human prion diseases. Missense mutations and expansions in the octapeptide repeat region of the gene are responsible for familial forms of prion disease. Five different mutations of the PRNP gene have been linked genetically to heritable prion disease.
Although phenotypes may vary dramatically within families, specific phenotypes tend to be observed with certain mutations. A clinical phenotype indistinguishable from typical sCJD is usually seen with substitutions at codons 180, 183, 200, 208, 210, and 232. Substitutions at codons 102, 105, 117, 198, and 217 are associated with the GSS variant of prion disease. The normal human PrP sequence contains five repeats of an eight-amino-acid sequence. Insertions from two to nine extra octarepeats frequently cause variable phenotypes ranging from a condition indistinguishable from sCJD to a slowly progressive dementing illness of many years in duration to an early-age-of-onset disorder that is similar to AD. A mutation at codon 178 resulting in substitution of asparagine for aspartic acid produces FFI if a methionine is encoded at the polymorphic residue 129 on the same allele. Typical CJD is seen if the D178N mutation occurs with a valine encoded at position 129 of the same allele.
HUMAN PRNP GENE POLYMORPHISMS
Polymorphisms influence the susceptibility to sporadic, inherited, and infectious forms of prion disease. The methionine/valine polymorphism at position 129 not only modulates the age of onset of some inherited prion diseases but can also determine the clinical phenotype. The finding that homozygosity at codon 129 predisposes to sCJD supports a model of prion production that favors PrP interactions between homologous proteins.
Substitution of the basic residue lysine at position 218 in mouse PrP produced dominant-negative inhibition of prion replication in transgenic mice. This same lysine at position 219 in human PrP has been found in 12% of the Japanese population, and this group appears to be resistant to prion disease. Dominant-negative inhibition of prion replication was also found with substitution of the basic residue arginine at position 171; sheep with arginine were resistant to scrapie prions but were susceptible to BSE prions that were inoculated intracerebrally.
INFECTIOUS PrP PRION DISEASES
IATROGENIC CJD
Accidental transmission of CJD to humans appears to have occurred with corneal transplantation, contaminated electroencephalogram (EEG) electrode implantation, and surgical procedures. Corneas from donors with unsuspected CJD have been transplanted to apparently healthy recipients who developed CJD after variable incubation periods. The same improperly decontaminated EEG electrodes that caused CJD in two young patients with intractable epilepsy caused CJD in a chimpanzee 18 months after their experimental implantation.
Surgical procedures may have resulted in accidental inoculation of patients with prions, presumably because some instrument or apparatus in the operating theater became contaminated when a CJD patient underwent surgery. Although the epidemiology of these studies is highly suggestive, no proof for such episodes exists.
Dura Mater Grafts More than 160 cases of CJD after implantation of dura mater grafts have been recorded. All of the grafts appear to have been acquired from a single manufacturer whose preparative procedures were inadequate to inactivate human prions. One case of CJD occurred after repair of an eardrum perforation with a pericardium graft.
Human Growth Hormone and Pituitary Gonadotropin Therapy The transmission of CJD prions from contaminated human growth hormone (hGH) preparations derived from human pituitaries has been responsible for fatal cerebellar disorders with dementia in >180 patients ranging in age from 10 to 41 years. These patients received injections of hGH every 2–4 days for 4–12 years. If it is thought that these patients developed CJD from injections of prion-contaminated hGH preparations, the possible incubation periods range from 4 to 30 years. Only recombinant hGH is now used therapeutically so that possible contamination with prions is no longer an issue. Four cases of CJD have occurred in women receiving human pituitary gonadotropin.
VARIANT CJD
The restricted geographic occurrence and chronology of vCJD raised the possibility that BSE prions had been transmitted to humans through the consumption of tainted beef. More than 190 cases of vCJD have occurred, with >90% of these in Britain. vCJD has also been reported in people either living in or originating from France, Ireland, Italy, Netherlands, Portugal, Spain, Saudi Arabia, United States, Canada, and Japan.
The steady decline in the number of vCJD cases over the past decade argues that there will not be a prion disease epidemic in Europe, similar to those seen for BSE and kuru. What is certain is that prion-tainted meat should be prevented from entering the human food supply.
The most compelling evidence that vCJD is caused by BSE prions was obtained from experiments in mice expressing the bovine PrP transgene. Both BSE and vCJD prions were efficiently transmitted to these transgenic mice and with similar incubation periods. In contrast to sCJD prions, vCJD prions did not transmit disease efficiently to mice expressing a chimeric human-mouse PrP transgene. Earlier studies with nontransgenic mice suggested that vCJD and BSE might be derived from the same source because both inocula transmitted disease with similar but very long incubation periods.
Attempts to determine the origin of BSE and vCJD prions have relied on passaging studies in mice, some of which are described above, as well as studies of the conformation and glycosylation of PrPSc. One scenario suggests that a particular conformation of bovine PrPSc was selected for heat resistance during the rendering process and was then reselected multiple times as cattle infected by ingesting prion-contaminated meat and bone meal (MBM) were slaughtered and their offal rendered into more MBM. Variant CJD cases have virtually disappeared with protection of the beef supply in Europe.
NEUROPATHOLOGY
Frequently the brains of patients with CJD have no recognizable abnormalities on gross examination. Patients who survive for several years have variable degrees of cerebral atrophy.
On light microscopy, the pathologic hallmarks of CJD are spongiform degeneration and astrocytic gliosis. The lack of an inflammatory response in CJD and other prion diseases is an important pathologic feature of these degenerative disorders. Spongiform degeneration is characterized by many 1- to 5-μm vacuoles in the neuropil between nerve cell bodies. Generally the spongiform changes occur in the cerebral cortex, putamen, caudate nucleus, thalamus, and molecular layer of the cerebellum. Astrocytic gliosis is a constant but nonspecific feature of prion diseases. Widespread proliferation of fibrous astrocytes is found throughout the gray matter of brains infected with CJD prions. Astrocytic processes filled with glial filaments form extensive networks.
Amyloid plaques have been found in ~10% of CJD cases. Purified CJD prions from humans and animals exhibit the ultrastructural and histochemical characteristics of amyloid when treated with detergents during limited proteolysis. In first passage from some human Japanese CJD cases, amyloid plaques have been found in mouse brains. These plaques stain with antibodies raised against PrP.
The amyloid plaques of GSS disease are morphologically distinct from those seen in kuru or scrapie. GSS plaques consist of a central dense core of amyloid surrounded by smaller globules of amyloid. Ultrastructurally, they consist of a radiating fibrillar network of amyloid fibrils, with scant or no neuritic degeneration. The plaques can be distributed throughout the brain but are most frequently found in the cerebellum. They are often located adjacent to blood vessels. Congophilic angiopathy has been noted in some cases of GSS disease.
In vCJD, a characteristic feature is the presence of “florid plaques.” These are composed of a central core of PrP amyloid, surrounded by vacuoles in a pattern suggesting petals on a flower.
CLINICAL FEATURES
Nonspecific prodromal symptoms occur in approximately a third of patients with CJD and may include fatigue, sleep disturbance, weight loss, headache, anxiety, vertigo, malaise, and ill-defined pain. Most patients with CJD present with deficits in higher cortical function. Similarly, psychiatric symptoms, such as depression, psychosis, and visual hallucinations, are often the defining features of the illness. These deficits almost always progress over weeks or months to a state of profound dementia characterized by memory loss, impaired judgment, and a decline in virtually all aspects of intellectual function. A few patients present with either visual impairment or cerebellar gait and coordination deficits. Frequently the cerebellar deficits are rapidly followed by progressive dementia. Visual problems often begin with blurred vision and diminished acuity, rapidly followed by dementia.
Other symptoms and signs include extrapyramidal dysfunction manifested as rigidity, masklike facies, or (less commonly) choreoathetoid movements; pyramidal signs (usually mild); seizures (usually major motor) and, less commonly, hypoesthesia; supranuclear gaze palsy; optic atrophy; and vegetative signs such as changes in weight, temperature, sweating, or menstruation.
Myoclonus Most patients (~90%) with CJD exhibit myoclonus that appears at various times throughout the illness. Unlike other involuntary movements, myoclonus persists during sleep. Startle myoclonus elicited by loud sounds or bright lights is frequent. It is important to stress that myoclonus is neither specific nor confined to CJD and tends to occur later in the course of CJD. Dementia with myoclonus can also be due to AD (Chap. 448), dementia with Lewy bodies (Chap. 448), corticobasal degeneration (Chap. 448) cryptococcal encephalitis (Chap. 239), or the myoclonic epilepsy disorder Unverricht-Lundborg disease (Chap. 445).
Clinical Course In documented cases of accidental transmission of CJD to humans, an incubation period of 1.5–2 years preceded the development of clinical disease. In other cases, incubation periods of up to 40 years have been suggested. Most patients with CJD live 6–12 months after the onset of clinical signs and symptoms, whereas some live for up to 5 years.
DIAGNOSIS
The constellation of dementia, myoclonus, and periodic electrical bursts in an afebrile 60-year-old patient generally indicates CJD. Clinical abnormalities in CJD are confined to the CNS. Fever, elevated sedimentation rate, leukocytosis in blood, or a pleocytosis in cerebrospinal fluid (CSF) should alert the physician to another etiology to explain the patient’s CNS dysfunction, although there are rare cases of CJD in which mild CSF pleocytosis is observed.
Variations in the typical course appear in inherited and transmitted forms of the disease. fCJD has an earlier mean age of onset than sCJD. In GSS disease, ataxia is usually a prominent and presenting feature, with dementia occurring late in the disease course. GSS disease presents earlier than CJD (mean age 43 years) and is typically more slowly progressive than CJD; death usually occurs within 5 years of onset. FFI is characterized by insomnia and dysautonomia; dementia occurs only in the terminal phase of the illness. Rare sporadic cases have been identified. vCJD has an unusual clinical course, with a prominent psychiatric prodrome that may include visual hallucinations and early ataxia, whereas frank dementia is usually a late sign of vCJD.
DIFFERENTIAL DIAGNOSIS
Many conditions mimic CJD. Dementia with Lewy bodies (Chap. 448) is the most common disorder to be mistaken for CJD. It can present in a subacute fashion with delirium, myoclonus, and extrapyramidal features. Other neurodegenerative disorders (Chap. 448) to consider include AD, FTD, corticobasal degeneration, progressive supranuclear palsy, ceroid lipofuscinosis, and myoclonic epilepsy with Lafora bodies (Chap. 445). The absence of abnormalities on diffusion-weighted and fluid-attenuated inversion recovery (FLAIR) magnetic resonance imaging (MRI) will almost always distinguish these dementing conditions from CJD.
Hashimoto’s encephalopathy, which presents as a subacute progressive encephalopathy with myoclonus and periodic triphasic complexes on the EEG, should be excluded in every case of suspected CJD. It is diagnosed by the finding of high titers of antithyroglobulin or antithyroid peroxidase (antimicrosomal) antibodies in the blood and improves with glucocorticoid therapy. Unlike CJD, fluctuations in severity typically occur in Hashimoto’s encephalopathy.
Intracranial vasculitides (Chap. 385) may produce nearly all of the symptoms and signs associated with CJD, sometimes without systemic abnormalities. Myoclonus is exceptional with cerebral vasculitis, but focal seizures may confuse the picture. Prominent headache, absence of myoclonus, stepwise change in deficits, abnormal CSF, and focal white matter changes on MRI or angiographic abnormalities all favor vasculitis.
Paraneoplastic conditions (Chap. 122), particularly limbic encephalitis and cortical encephalitis, can also mimic CJD. In many of these patients, dementia appears prior to the diagnosis of a tumor, and in some, no tumor is ever found. Detection of the paraneoplastic antibodies is often the only way to distinguish these cases from CJD.
Other diseases that can simulate CJD include neurosyphilis (Chap. 206), AIDS dementia complex (Chap. 226), progressive multifocal leukoencephalopathy (Chap. 164), subacute sclerosing panencephalitis, progressive rubella panencephalitis, herpes simplex encephalitis (Chap. 164), diffuse intracranial tumor (gliomatosis cerebri; Chap. 118), anoxic encephalopathy, dialysis dementia, uremia, hepatic encephalopathy, voltage-gated potassium channel (VGkC) autoimmune encephalopathy, and lithium or bismuth intoxication.
LABORATORY TESTS
The only specific diagnostic tests for CJD and other human prion diseases measure PrPSc. The most widely used method involves limited proteolysis that generates PrP 27-30, which is detected by immunoassay after denaturation. The conformation-dependent immunoassay (CDI) is based on immunoreactive epitopes that are exposed in PrPC but buried in PrPSc. In humans, the diagnosis of CJD can be established by brain biopsy if PrPSc is detected. If no attempt is made to measure PrPSc, but the constellation of pathologic changes frequently found in CJD is seen in a brain biopsy, then the diagnosis is reasonably secure (see “Neuropathology,” above). The high sensitivity and specificity of cortical ribboning and basal ganglia hyperintensity on FLAIR and diffusion-weighted MRI for the diagnosis of CJD have greatly diminished the need for brain biopsy in patients with suspected CJD. Because PrPSc is not uniformly distributed throughout the CNS, the apparent absence of PrPSc in a limited sample such as a biopsy does not rule out prion disease. At autopsy, sufficient brain samples should be taken for both PrPSc immunoassay, preferably by CDI, and immunohistochemistry of tissue sections.
To establish the diagnosis of either sCJD or familial prion disease, sequencing the PRNP gene must be performed. Finding the wild-type PRNP gene sequence permits the diagnosis of sCJD if there is no history to suggest infection from an exogenous source of prions. The identification of a mutation in the PRNP gene sequence that encodes a nonconservative amino acid substitution argues for familial prion disease.
CT may be normal or show cortical atrophy. MRI is valuable for distinguishing sCJD from most other conditions. On FLAIR sequences and diffusion-weighted imaging, ~90% of patients show increased intensity in the basal ganglia and cortical ribboning (Fig. 453e-3). This pattern is not seen with other neurodegenerative disorders but has been seen infrequently with viral encephalitis, paraneoplastic syndromes, or seizures. When the typical MRI pattern is present, in the proper clinical setting, diagnosis is facilitated. However, some cases of sCJD do not show this typical pattern, and other early diagnostic approaches are still needed.
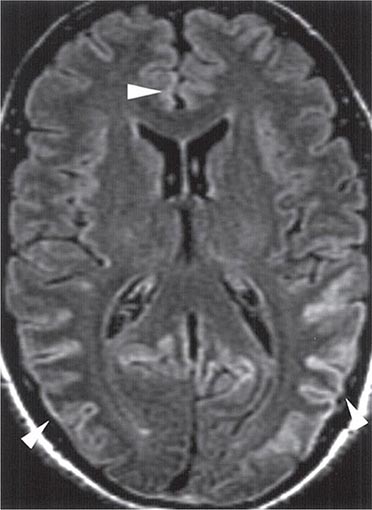
FIGURE 453e-3 T2-weighted (fluid-attenuated inversion recovery) magnetic resonance imaging showing hyperintensity in the cortex in a patient with sporadic CJD. This so-called “cortical ribboning” along with increased intensity in the basal ganglia on T2- or diffusion-weighted imaging can aid in the diagnosis of Creutzfeldt-Jakob disease.
CSF is nearly always normal but may show protein elevation and, rarely, mild pleocytosis. Although the stress protein 14-3-3 is elevated in the CSF of some patients with CJD, similar elevations of 14-3-3 are found in patients with other disorders; thus this elevation is not specific. Similarly, elevations of CSF neuron-specific enolase and tau occur in CJD but lack specificity for diagnosis.
The EEG is often useful in the diagnosis of CJD, although only approximately 60% of individuals show the typical pattern. During the early phase of CJD, the EEG is usually normal or shows only scattered theta activity. In most advanced cases, repetitive, high-voltage, triphasic, and polyphasic sharp discharges are seen, but in many cases their presence is transient. The presence of these stereotyped periodic bursts of <200 ms in duration, occurring every 1–2 s, makes the diagnosis of CJD very likely. These discharges are frequently but not always symmetric; there may be a one-sided predominance in amplitude. As CJD progresses, normal background rhythms become fragmentary and slower.
CARE OF CJD PATIENTS
Although CJD should not be considered either contagious or communicable, it is transmissible. The risk of accidental inoculation by aerosols is very small; nonetheless, procedures producing aerosols should be performed in certified biosafety cabinets. Biosafety level 2 practices, containment equipment, and facilities are recommended by the Centers for Disease Control and Prevention and the National Institutes of Health. The primary problem in caring for patients with CJD is the inadvertent infection of health care workers by needle and stab wounds. Electroencephalographic and electromyographic needles should not be reused after studies on patients with CJD have been performed.
There is no reason for pathologists or other morgue employees to resist performing autopsies on patients whose clinical diagnosis was CJD. Standard microbiologic practices outlined here, along with specific recommendations for decontamination, seem to be adequate precautions for the care of patients with CJD and the handling of infected specimens.
DECONTAMINATION OF CJD PRIONS
Prions are extremely resistant to common inactivation procedures, and there is some disagreement about the optimal conditions for sterilization. Some investigators recommend treating CJD-contaminated materials once with 1 N NaOH at room temperature, but we believe this procedure may be inadequate for sterilization. Autoclaving at 134°C for 5 h or treatment with 2 N NaOH for several hours is recommended for sterilization of prions. The term sterilization implies complete destruction of prions; any residual infectivity can be hazardous. Recent studies show that sCJD prions bound to stainless steel surfaces are resistant to inactivation by autoclaving at 134°C for 2 h; exposure of bound prions to an acidic detergent solution prior to autoclaving rendered prions susceptible to inactivation.
PREVENTION AND THERAPEUTICS
There is no known effective therapy for preventing or treating CJD. The finding that phenothiazines and acridines inhibit PrPSc formation in cultured cells led to clinical studies of quinacrine in CJD patients. Unfortunately, quinacrine failed to slow the rate of cognitive decline in CJD, possibly because therapeutic concentrations in the brain were not achieved. Although inhibition of the P-glycoprotein (Pgp) transport system resulted in substantially increased quinacrine levels in the brains of mice, the prion incubation times were not extended by treatment with the drug. Whether such an approach can be used to treat CJD remains to be established.
Like the acridines, anti-PrP antibodies have been shown to eliminate PrPSc from cultured cells. Additionally, such antibodies in mice, either administered by injection or produced from a transgene, have been shown to prevent prion disease when prions are introduced by a peripheral route, such as intraperitoneal inoculation. Unfortunately, the antibodies were ineffective in mice inoculated intracerebrally with prions. Several drugs, including pentosan polysulfate as well as porphyrin and phenylhydrazine derivatives, delay the onset of disease in animals inoculated intracerebrally with prions if the drugs are given intracerebrally beginning soon after inoculation.
DIFFERENT PRIONS CAUSING OTHER NEURODEGENERATIVE DISEASES
There is a rapidly expanding body of literature demonstrating that in addition to PrP, other proteins including amyloid beta (Aβ), tau, α-synuclein, and huntingtin can all becomes prions (Chap. 444e). Experimental studies have shown that transgenic mice expressing mutant amyloid precursor protein (APP) develop amyloid plaques containing fibrils composed of the Aβ peptide approximately a year after inoculation with synthetic Aβ peptides polymerized into amyloid fibrils or extracts prepared from the brains of patients with AD. Mutant tau aggregates in transgenic mice and cultured cells can trigger the aggregation of tau into fibrils that resemble those found in neurofibrillary tangles and Pick bodies. Such tangles have been found in AD, FTDs, Pick’s disease, and some cases of posttraumatic brain injury, all of which are likely to be caused by the prion isoforms of Aβ and/or tau.
In patients with advanced PD who received grafts of fetal substantia nigral neurons, Lewy bodies containing β-sheet–rich α-synuclein were identified in grafted cells approximately 10 years after transplantation, arguing for the axonal transport of misfolded α-synuclein crossing into grafted neurons, where it initiated aggregation of nascent α-synuclein into fibrils that coalesced into Lewy bodies. These findings combined with studies of multiple system atrophy (MSA) argue that the synucleinopathies are caused by prions. Brain homogenates from MSA patients injected into transgenic mice transmitted lethal neurodegeneration in approximately 3 months; moreover, recombinant synuclein injected into wild-type mice initiated the deposition of synuclein fibrils.
In summary, a wealth of evidence continues to accumulate arguing that proteins causing AD, PD, FTDs, ALS, and even HD acquire alternative conformations that become self-propagating. Each of these neurodegenerative diseases is caused by a different protein that undergoes a conformational transformation to become a prion. Prions explain many of the features that the neurodegenerative diseases have in common: (1) incidence increases with age, (2) steady progression over years, (3) spread from one region of the CNS to another, (4) protein deposits consisting of amyloid fibrils, and (5) late onset of inherited forms of the neurodegenerative diseases. Notably, amyloid plaques containing PrPSc are a nonobligatory feature of PrP prion disease in humans and animals. Furthermore, amyloid plaques in AD do not correlate with the level of dementia; however, the level of soluble (oligomeric) Aβ peptide does correlate with memory loss and other intellectual deficits.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


