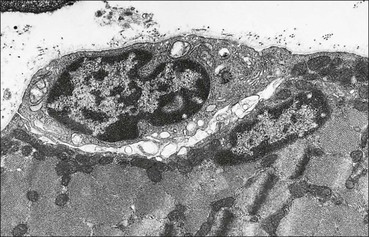
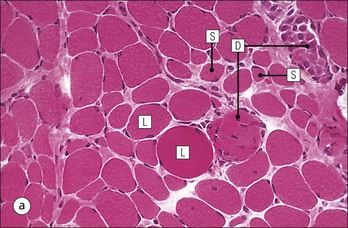
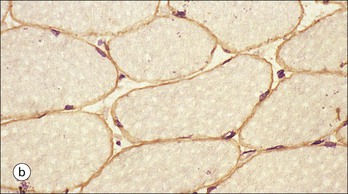
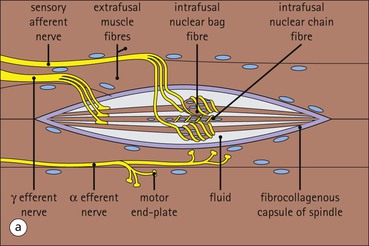
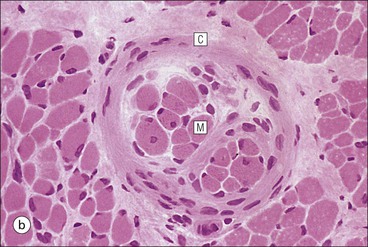
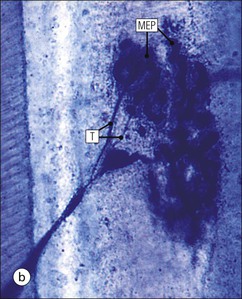
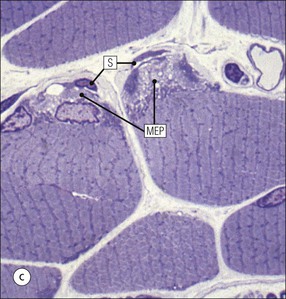
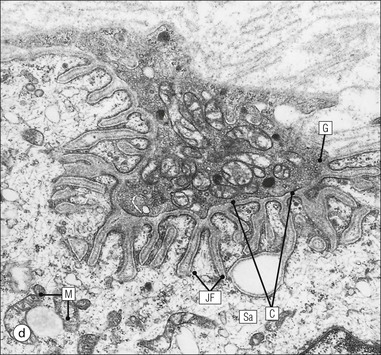
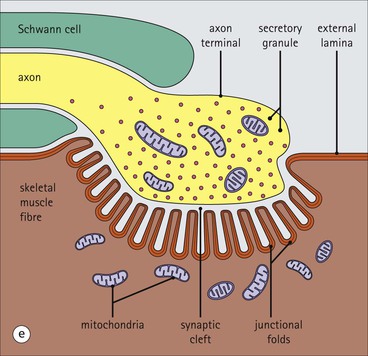
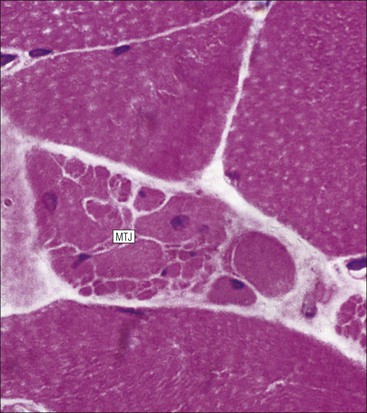
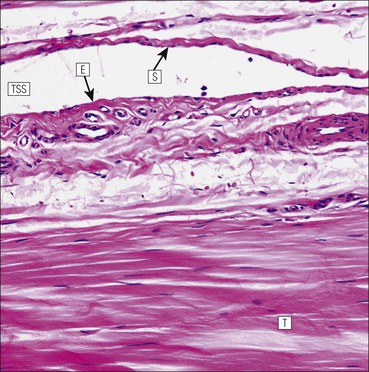
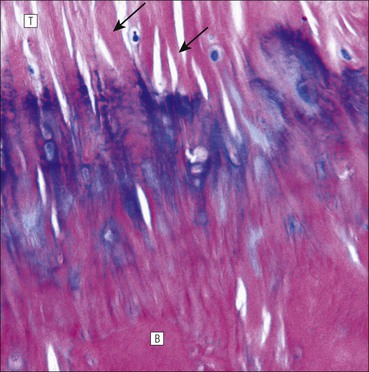
The musculoskeletal system, which provides mechanical support and permits movement, is composed of skeletal muscle, tendons, bones, joints and ligaments. Skeletal muscles act as contractile levers, and are connected to bone by tendons. Bones act as rigid levers, articulating with other bones through joints, which are kept in relationship by ligaments. A special characteristic of muscle, tendons and joints is the possession of a rich sensory nerve supply, which detects the position of the body and the velocity of movement. The integration of this sensory information by the central nervous system is vital for the musculoskeletal system to function normally. The main functional attribute of bone is its specialized extracellular matrix, which is hardened by the deposition of calcium enabling it to function as a rigid lever. The rigid, hard character of bone belies its importance as a metabolic reservoir of mineral salts and the fact that it is in a constant state of dynamic remodelling. Skeletal muscle is responsible for voluntary movement The histological characteristics of skeletal muscle cells and the structural basis of muscle contraction are described on pages 71–76. Individual muscle cells are arranged into large groups to form anatomically distinct muscles. These are characterized by: • An orderly alignment of the constituent cells to generate a directional force following contraction • Anchorage to other structures by highly organized fibrocollagenous support tissues • A rich blood supply reflecting high metabolic demands Embryologically, muscle fibres develop from mesenchymal tissues In embryogenesis, skeletal muscle develops from primitive mesenchymal tissues of the mesoderm. Small spindle-shaped or strap-shaped cells with a single nucleus and abundant pink-staining cytoplasm develop from primitive mesenchymal cells. These are skeletal muscle cell precursors, the rhabdomyoblasts (Fig. 13.1). Numerous individual rhabdomyoblasts then fuse to form multinucleate muscle fibres, the cells enlarging in size when they become connected to the nervous system and receive motor stimulation. Residual cells with the function of rhabdomyoblasts (i.e. capable of differentiating further into functional striated muscle cells) persist in adult skeletal muscle as satellite cells (see Fig. 13.6). Following muscle damage and loss of fibres, these cells proliferate and may produce new muscle cells. A muscle is composed of many muscle fibres A mature muscle fibre is composed of large numbers of myofibrils bounded by a cell membrane termed ‘the sarcolemma’ (see Figs 5.2, 13.2). Each skeletal muscle cell is typically extremely long (up to 10 cm in length), and it is therefore more usual to use the term ‘skeletal muscle fibre’ rather than cell. Many muscle fibres are arranged together to form a muscle (Fig. 13.3). Despite being elongated, individual muscle cells do not extend the full length of a muscle but are arranged in overlapping bundles, the force of contraction being transmitted through the arrangement of the support tissues. Different muscles are characterized by different physiological and metabolic properties The physiological and metabolic properties of individual skeletal muscles are determined by differences in the structure of their constituent muscle fibres. In both animals and man, it has been possible to define several subtypes of muscle fibre by macroscopic, physiological, biochemical and histochemical criteria, but there are marked interspecies variations. Histochemical staining for specific enzymes delineates several types of fibres and is a useful method for analysing muscle. Two main types of fibres are identified (types 1 and 2), and type 2 fibres can be subdivided into types 2A, 2B and 2C (Fig. 13.4). Type 2C fibres are thought to be a primitive form of type 2 fibre and, with appropriate innervation, are probably capable of developing into 2A or 2B fibres. Such histochemical staining is used routinely in the study of muscle pathology and allows the histological diagnosis of certain muscle diseases. Histochemical staining can also be correlated with other functional and biochemical attributes of muscle (Fig. 13.4c). Not all muscles have the same proportions of type 1 and type 2 fibres. In general, muscles with a role in maintaining posture (e.g. calf muscles) have a high proportion of type 1 fibres, whereas muscles used for short bursts of power have abundant type 2 fibres. Different individuals are genetically endowed with variable proportions of muscle fibre types in defined muscles, and this may limit athletic prowess at a particular sport; training does not affect the proportion of fibres in any given muscle, but does influence their size. Muscle has a rich blood supply because of the high energy demands of contraction Large arteries penetrate the epimysium and divide into small branches that run in the perimysial support tissues, forming perimysial arteries and veins. These branches terminate in a vast capillary network, which runs in the endomysium. Each muscle fibre is served by several capillary vessels. Normal adult skeletal muscle cells do not undergo cell division Any increased demands placed on a muscle, for example by weight training, result in increased muscle size because the cells themselves increase in size (i.e. hypertrophy). It is possible, however, for skeletal muscle cells to regrow because the pool of inactive stem cells in adult muscle called ‘satellite cells’ can be stimulated to divide following damage. These cells are not discernible by light microscopy, but may be seen ultrastructurally (Fig. 13.6). They are infrequent and appear as small spindle-shaped cells lying just beneath the external lamina of a muscle fibre. Diseases of Muscle Several diseases of muscle have been attributed to specific metabolic or structural abnormalities (Fig. 13.5a). Duchenne muscular dystrophy is the most common inherited muscle disease and characteristically affects male children. Such individuals become unable to stand unaided in early childhood and develop progressive muscle weakness, becoming wheelchair-bound by their mid-teens and typically dying in early adult life. The abnormality in Duchenne muscular dystrophy is due to a defect in the gene coding for a protein termed ‘dystrophin’ (Fig. 13.5b). This protein links the actin cytoskeleton of muscle to the external lamina; its absence leads to abnormal muscle fibre fragility. Fibres that have regenerated in adult life following damage to a muscle can be detected histologically because they commonly contain centrally placed nuclei, rather than the peripheral nuclei typical of normal fibres. Muscle contains stretch receptors Although there are no pain receptors in skeletal muscle, there are receptors sensitive to stretch, which function as part of a feedback system to maintain normal muscle tone (i.e. the spinal stretch reflex arc). Sensory fibres that provide information on the tension of skeletal muscle arise from two sources: • Encapsulated nerve endings responding to stretch in the tendon associated with a muscle • Spiral nerve endings (sensory afferent fibres) sensing stretch and tension of specialized muscle fibres contained in a special sense organ in muscle called the muscle spindle (Fig. 13.7). The muscle spindle sensory mechanism maintains normal tone and muscle coordination. Motor nerves to muscle terminate in motor endplates Large nerves, containing both motor and sensory axons, enter muscles by penetrating the epimysium and branch to form small nerves, which run in the perimysium. Perimysial nerves contain axons to fulfil both motor and sensory functions. The motor axons destined to innervate skeletal muscle (α efferent fibres) enter the endomysium as nerve twigs and branch to innervate several fibres. At the end of each twig, the axon becomes modified to form a motor endplate, and it is this that activates skeletal muscle contraction (Fig. 13.8). Activation of the motor axon causes the release of acetylcholine from its storage granules by exocytosis. Acetylcholine then diffuses across the gap between the axon and muscle fibre and interacts with specific membrane receptors to cause depolarization of the muscle fibre, which initiates contraction (see p. 73). The activity of secreted acetylcholine is rapidly curtailed by the activity of an enzyme called ‘acetylcholinesterase’, which is bound to the basement membrane investing the junctional folds. In addition to nerve fibres controlling voluntary movement, specialized motor axons (γ efferent fibres) innervate fibres in the muscle spindle. Myasthenia Gravis Myasthenia gravis is a disease caused by the formation of antibodies to the acetylcholine receptor on the sarcolemma in the junctional folds of the motor endplates. The antibodies bind to the acetylcholine receptors and thereby prevent released acetylcholine from interacting with the receptors and causing depolarization. Affected individuals develop tremendous muscle weakness manifest by fatiguability, inability to lift the arms, failure to maintain an upright posture of the head, and drooping of the eyelids. Treatment is by the administration of drugs (anticholinesterases) that inhibit the action of the enzyme acetylcholinesterase. This potentiates the action of released acetylcholine and allows it to bind to receptors not blocked by antibody. Myasthenia gravis is an autoimmune disease. In order to transmit the force of contraction, the cell surface at the end of the muscle fibres is adapted for attachment to highly organized fibrocollagenous support tissues, which anchor the muscle to other structures. Such anchorage may be provided by: Tendons attach some muscles to bone Muscles are connected to bone by dense compact collagenous tissue called ‘tendinous tissue’, which may be in the form of short sheets attaching to bone over a large area, often linear, or long cylindrical structures which connect some muscles to the bones of hands and fingers, and feet and toes. The largest and most distinct of the cylindrical type of tendons is the Achilles tendon, which connects the substantial muscle masses of the back of the calf with the bones at the back of the foot, involved in flexion and extension of the foot. A series of long cylindrical tendons run through the hands to the fingers. Tendinous tissue is composed of tightly packed, longitudinally running collagen fibres, with the elongated flattened muscle nuclei of inactive fibrocytes scattered between them (Fig. 13.9). In tendon, these fibrocytes are sometimes called ‘tenocytes’ or ‘tendinocytes’. Tendinous tissue is relatively acellular and has both low oxygen/nutrient requirements and a very limited blood supply. Because of this poor vascularity, tendons that have been damaged by trauma (usually due to acute overstretching, particularly in sporting injuries) are extremely slow to heal, e.g. Achilles tendon rupture, extensor tendonitis in the elbow (‘tennis elbow’). Tendons attach to skeletal muscles at specialized structures called myotendinous junctions The collagen fibres of tendons are firmly tethered to the muscle fibres of skeletal muscles at specialized areas called myotendinous insertions or junctions (Fig. 13.10). At these sites, individual muscle fibres develop a complex interdigitating surface, which is tightly anchored to the collagen fibres of the intramuscular portion of the end of the tendon. Many tendons lie within fibrocollagenous tendon sheaths Where the long cylindrical type of tendon has to run over the surface of bone it must be protected from damage due to repeated friction. An example is the long flexor tendons of the hand running to the fingers. This protection from friction damage is achieved by the tendon having a thin outer fibrous capsule sometimes called the epitendineum, coated on its outer surface by flat mesothelial cells, which are synovial in type and secrete small amounts of lubricating fluid similar to that which lubricates synovial joints (see p. 260). Such tendons are surrounded by a thin collagenous sheath which is lined internally by similar synovial-type mesothelial cells, also secreting lubricating fluid. This structure is called the tendon sheath, and the synovium related to the epitendineum and the tendon sheath is called tenosynovium. Thus, the tendon can slide effortlessly and without friction within a lubricated sheath (Fig. 13.11). Despite these preventive measures, repeated overuse of some tendons (particularly in the hands) can lead to structural damage to both epitendineum and tendon sheath, which is manifest as a painful condition localized to the affected tendon (‘tendonitis’). Various forms of tendonitis, particularly those affecting the hands and arms, are often regarded as components of so-called ‘repetitive strain injury’. Tendon is attached to bone by fibres which pass through the periosteum and penetrate into the bone At the attachment to bone, the type I collagen fibres of a tendon merge with the type I collagen of the periosteum and send fibres that penetrate into the bone (Fig. 13.12). These are called Sharpey’s fibres. The original tendon collagen is probably incorporated into bone when the periosteum makes new bone, initially in bone development in intrauterine life, and is continually reinforced in periods of high periosteal activity during bone growth in childhood. Maintenance of the tendon at the site of the bone attachment is the responsibility of the collagen-synthesizing fibroblasts of the periosteum. These cells have the capacity to convert into chondroprogenitor cells (chondroblast precursors) or osteoprogenitor cells (osteoblast precursors, see Fig. 13.16). For this reason, the tendon close to its attachment to the bone may contain small islands of cartilage or bone. Other important structures in the musculoskeletal system have a structure similar to that of tendon Ligaments join one bone to another (see Fig. 13.29). Like tendons, they are composed of tightly packed collagen fibres running predominantly in the same direction, but less regularly than in long cylindrical tendons, with inactive elongated fibrocytes lying between the collagen bundles. In some areas there is a need for the ligaments to have elasticity, for example in the prominent and extensive ligaments that help to bind the bones of the vertebral column together (e.g. ligamentum flavum). In such areas the ligament contains abundant elastic fibres which also run, regularly spaced, parallel with the collagen fibres; these are sometimes called elastic ligaments. Aponeuroses are flat sheets, often extensive, of tendon-like compact fibrocollagenous tissue, which may connect muscle to muscle (e.g. in the anterior abdominal wall), muscle to bone at an insertion, or muscle to long cylindrical types of tendon. Thus, many aponeuroses can be considered to be part of the tendinous system. Because the muscle fibres are attached to an aponeurosis in different directions, the component collagen of the aponeurosis is arranged in layers that run in different directions. Large aponeuroses are found in the palms and soles. Bone is a highly specialized support tissue which is characterized by its rigidity and hardness. Its four main functions are to provide mechanical support (e.g. ribs), to permit locomotion (e.g. long bones), to provide protection (e.g. skull) and to act as a metabolic reservoir of mineral salts. Bone is composed of: • Support cells (osteoblasts and osteocytes) • A non-mineral matrix of collagen and glycosaminoglycans (osteoid) • Inorganic mineral salts deposited within the matrix Bone contains a specialized extracellular matrix material called osteoid Osteoid is a collagenous support tissue of type I collagen embedded in a glycosaminoglycan gel containing specific glycoproteins (e.g. osteocalcin) that strongly bind calcium. Deposition of mineral salts in the osteoid gives bone its characteristic rigidity and functional strength. Osteoid is synthesized by specialized support cells called ‘osteoblasts’, which have the same collagen synthesis capacity as fibroblasts (see p. 63). Formed calcified bone can be eroded by osteoclasts, modified multinucleate phagocytic cells belonging to the monocyte/macrophage family (see p. 128). Coordinated activity of osteoclasts (eroding formed bone) and osteoblasts (synthesized new osteoid) is important in maintaining bone structure, and any structural modifications necessary to meet increased or altered demands. This is called ‘bone remodelling’. Bone is constantly being remodelled Bone is a dynamic tissue, being continually formed and destroyed under the control of hormonal and physical factors. This constant activity permits the process of modelling (i.e. modification of the bone architecture to meet physical stresses). Bone turnover is normally low in adults, but in babies and children it is high to allow the growth and active remodelling required to cope with new demands, for example with the onset of walking.
Musculoskeletal System
Introduction
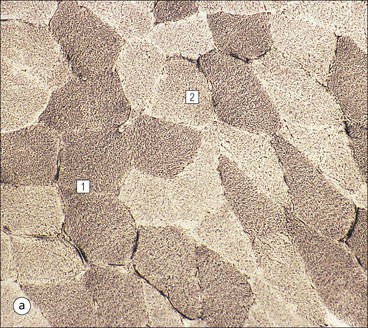
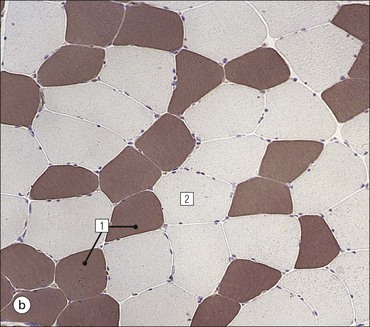
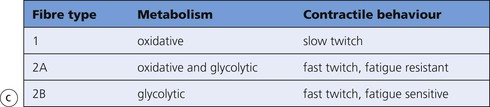
Skeletal Muscle
 Clinical Example
Clinical Example
 Clinical Example
Clinical Example
Muscle Attachments
Bone
Introduction
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Musculoskeletal System
Chapter 13
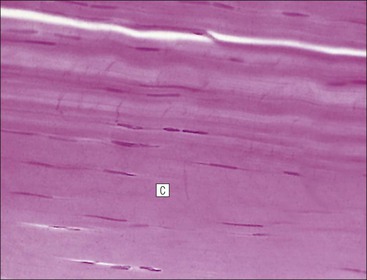
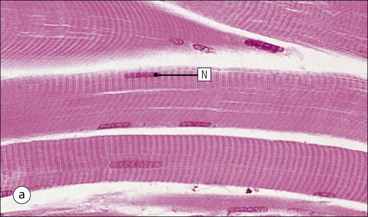
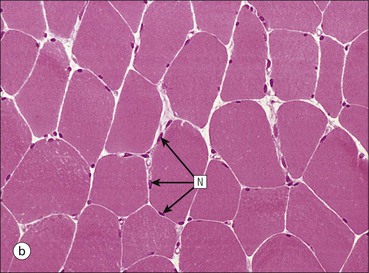
FIGURE 13.2 Skeletal muscle fibres. (a) Micrograph of skeletal muscle fibres in a longitudinal section showing prominent cross-striations. The dark bands are termed ‘A bands’ (anisotropic, i.e. birefringent in polarized light), and the light bands are termed ‘I bands’ (isotropic, i.e. no interference with polarized light). These bands correspond to the arrangement of thick and thin filaments in myofibrils (see Fig. 5.2). Nuclei (N) appear as elongated structures just beneath the cell membrane, and each fibre contains many nuclei. (b) Micrograph of skeletal muscle fibres in a transverse frozen section showing roughly hexagonal profiles with flattened sides where they are compressed by adjacent fibres. Nuclei (N) appear as small circular profiles at the periphery of each fibre. Individual myofibrils are generally not identifiable by light microscopy.
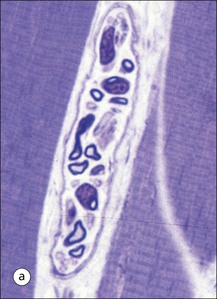
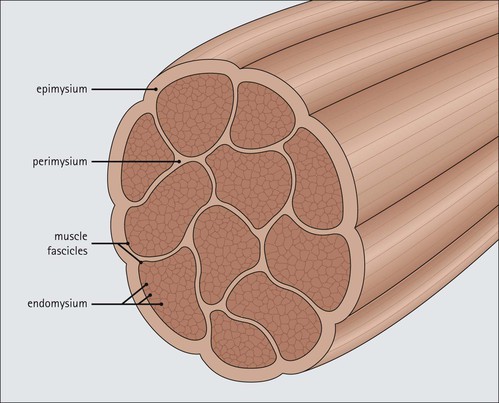
FIGURE 13.3 Organization of muscle fibres into muscle. Individual muscle fibres (see Fig. 5.1) are surrounded by endomysium, which is composed of sheets of external lamina identical to basement membrane (see p. 60). Endomysium secures the muscle fibres to each other and contains both capillary blood vessels and individual nerve axons (see Chapter 6). Clusters of muscle fibres are held together by fine sheets of fibrocollagenous support tissue (perimysium) to form fascicles. Blood vessels, lymphatic vessels and nerves run in the endomysial support tissues. An anatomically defined muscle is composed of many fascicles, which are surrounded externally by a thick layer of fibrocollagenous support tissue, the epimysium.