12 Locomotor system
ANATOMY
VERTEBRAL COLUMN
The vertebral column is made up of 32–34 vertebra:
The vertebral column as a whole is:
Viewed from its lateral aspect the column has a series of curves:
From the functional standpoint, the occipitoatlantoaxial region may be considered separately from the remainder of the mobile vertebral column (C3–L5). The majority of cervical rotation occurs at atlantoaxial level, while nodding movements and fine positioning (tilting) of the head on the neck occur mainly between atlas and occiput. In the rest of the mobile column, the final range of movement results from the summation of small movements at individual intervertebral levels (‘motion segments’). The regions with the most movement, and hence the least stability, are the junctions of the more ‘fixed’ levels (thoracic and sacral) with the more mobile levels (cervical and lumbar). The commonest forms of the unstable spinal injury occur at these junctional regions. At mid- and lower cervical levels and in the lumbar region the configuration of the facet joints, the arrangement of the muscles and the topography of the neighbouring soft tissues allow (forward) flexion, extension and lateral flexion (side-bending) movements which are largely prevented at thoracic level by the presence of the ribcage. Axial rotation (about the vertical axis of thecolumn) occurs maximally at thoracic level.
Osteology
Basic vertebral pattern
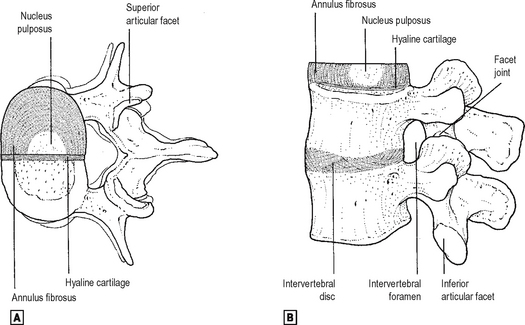
The basic vertebral pattern consists of a body and a neural arch surrounding a vertebral canal (Fig. 12.1).
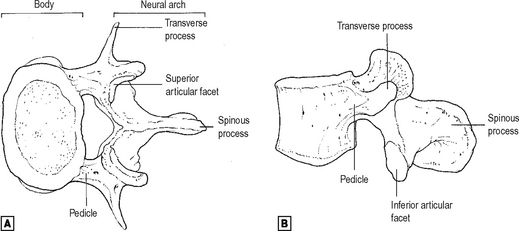
 superior view.
superior view.  lateral view.
lateral view.Special additional features at specific vertebral levels
Vertebrae from individual regions have distinguishing features.
Cervical (Fig. 12.2)
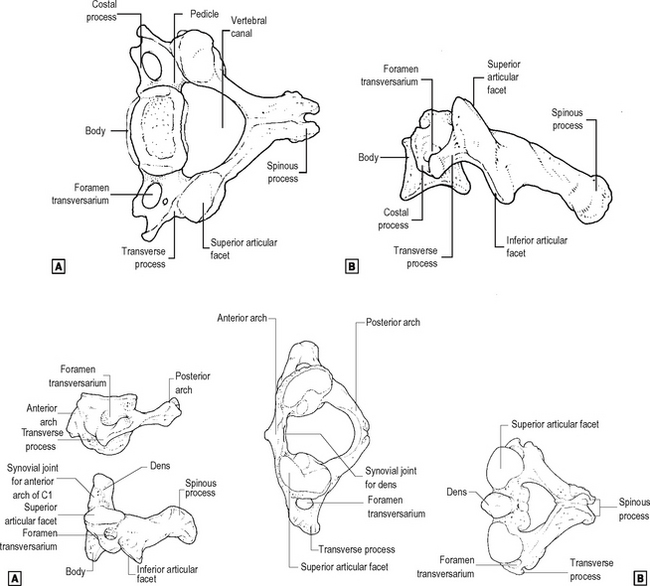
 superior view.
superior view.  lateral view. (2) The atlas and axis, cervical vertebrae 1 and 2.
lateral view. (2) The atlas and axis, cervical vertebrae 1 and 2.  lateral views.
lateral views.  superior views.
superior views.Thoracic (Fig. 12.3)
These are characterized by demifacets on the sides of the body of the vertebra for articulation with the heads of the ribs. They are characterized also by facets on their transverse processes for the rib tubercles apart from the lower two thoracic vertebrae. They are also characterized by long and downward sloping spines and their articular facets are relatively vertical.
 superior view.
superior view.  lateral view.
lateral view.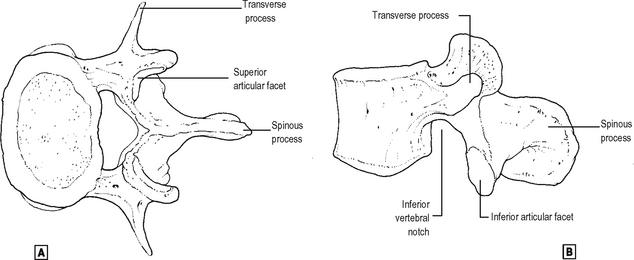
 superior view.
superior view. lateral view.
lateral view.Arthrology
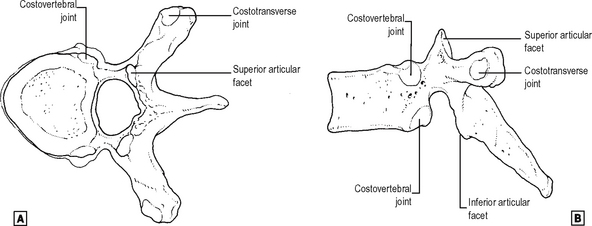
The mobile vertebral column can be considered functionally as a series of linked pairs of vertebrae. Each pair of vertebrae and the linkage between its members constitute a motion segment. These linkages include joints, ligaments and muscles. Although movement between adjacent vertebra is slight, the additive affect is considerable. Movement particularly occurs at the cervico-thoracic and thoraco-lumbar junctions and hence these are the two common sites of vertebral injury. A typical intervertebral joint is shown in Fig. 12.5.
 superior view.
superior view.  lateral view.
lateral view.The joints between atlas, axis and the skull are shown in Fig. 12.6.
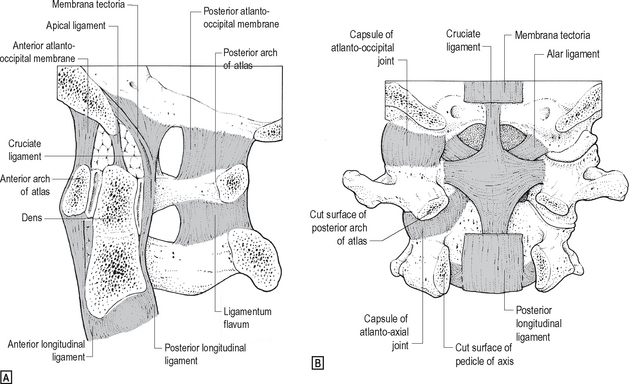
 midline sagittal section.
midline sagittal section.  posterior surface. The posterior arch of the atlas and the vertebral arch of the axis have been removed.
posterior surface. The posterior arch of the atlas and the vertebral arch of the axis have been removed.Movements and muscles
Transitional zones
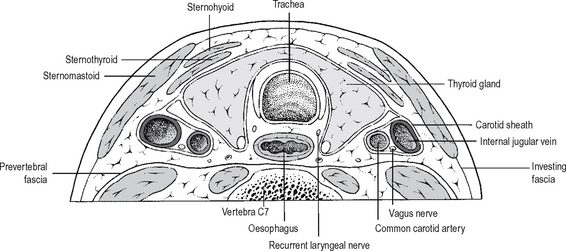
In the neck, the interface is demarcated by the prevertebral fascia: the visceral ‘column’ of the neck moves actively and passively on the musculoskeletal ‘column’ in the plane of this fascia. The fascial anatomy of the neck is of great surgical importance, and is best seen in transverse (axial) sections (Fig. 12.7). The prevertebral fascia of the neck extends inferiorly as the endothoracic fascia and subsequently the retroperitoneal fasciaof the abdomen.
Neurovascular patterns and relationships: the ‘holes’ and what they contain
The vertebral canal contains the spinal cord and cauda equina in their meningeal coverings, an incomplete epidural layer of fat containing the internal venous plexus, and some tenuous connective tissue connecting the dural tube or theca with the posterior longitudinal ligament. The most lateral parts of the canal contain the roots of the spinal nerves in their dural sheaths, within which each pair of dorsal and ventral roots joins, immediately lateral to the dorsal root ganglion. The epidural ‘space’ is only ‘potential’ until defined by air or liquid let into it as a result of pathology or medical intervention (e.g. epidural injection, epidural anaesthesia, laminectomy). The cross-sectional size and shape of the vertebral canal varies with vertebral level, as does the proportion of its cross-sectional area occupied by the spinal cord and its coverings. The normal canal is large in the upper cervical and lower lumbar regions, and smallest in the midthoracic region.
Each typical intervertebral foramen (Fig. 12.5) is bounded superiorly and inferiorly by the pedicles, anteriorly mainly by the intervertebral disc and posteriorly mainly by the facet joint. The foramen contains the foraminal segment of the exiting ‘nerve root’ (spinal nerve), together with the radicular vessels and finerecurrent meningeal (sinuvertebral) nerves. Owing to the varying obliquity and length of the nerve roots, which run increasingly distally as the spine is descended, the dorsal root ganglia may lie in the lateral zone of the vertebral canal or in the intervertebral foramen.
The main clinically relevant variations in the neurovascular patterns concern the effect of vertebral segmentation anomalies on the number and position of nerve roots (spinal nerves) exiting the vertebral column. Such segmentation anomalies are not uncommon, and chiefly affect the lumbosacral region. The lumbar spine may be shortened to four vertebrae (‘sacralised L5’), or lengthened to six (‘lumbarised S1’), with accompanying rearrangement of the nerve roots leading to possible confusion in the clinical assessment of neurological signs in the lower limbs.
In spinal injuries, the spinal cord, the nerve roots and the spinal nerves may be involved separately or together. The site of injury may be within the vertebral canal or in the intervertebral foramen and ‘root canal’. Both cord and roots may be involved at any level above L1, where the cord stops in an adult. Below L1 only the roots of the cauda equina or the spinal nerves can be affected. The motor signs in spinal injury may be those of an upper motor neuron lesion, a lower motor neuron lesion, or a mixed picture may be seen. The whole spectrum of severity of nerve injury is possible, including the effects of acute or chronic nerve compression. The injury may be open (rarely) or closed, and the cause may be spinal fracture-dislocation, disc prolapse, or bony compression (spinal stenosis, which can occur at one or several spinal levels and which may affect the whole vertebral canal or only its lateral recesses). Clinical examination should include testing for both motor and sensory signs of root lesions: familiarity with dermatomes andmuscle ‘root values’ is essential (Chapter 8).
PECTORAL GIRDLE
Osteology
Clavicle
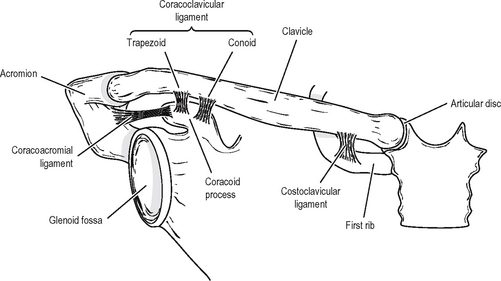
This is one of the most frequently fractured bones, and has important muscle and ligament attachments and soft-tissue relations. It is subcutaneous throughout its length, but the overlying skin is very mobile, and open fracture is uncommon. The lateral half of the bone is concave forwards and flattened, with prominent anterior and posterior borders, while the medial half is convex forwards and almost cylindrical. The inferior surface is roughened laterally for the attachment of the coracoclavicular ligament and medially for that of the costoclavicular ligament (Fig. 12.8). These ligaments are very strong: the bone always fractures between their attachments, in the segment on the inferior surface of which the subclavius muscle is attached. Deltoid and pectoralis major are attached to the anterior surface, while trapezius attaches posterolaterally. The clavicular head of sternocleidomastoid attaches superomedially. The most important soft-tissue relations lie posteriorly, particularly at the medial end where the subclavian and brachiocephalic vessels are near. The divisions of the brachial plexus lie behind the medial two-thirds of the bone, separated from it by anoften surprisingly large suprascapular vein. The clavicle is crossed anteriorly and subcutaneously by the palpable supraclavicular nerves from the cervical plexus.
Scapula (Fig. 12.9)
The scapula lies posterolaterally over the second to seventh ribs. It consists of a flat, thin triangular body whose lateral border is thickened for force transmission and to act as a lever for the action of the attached teres muscles. A notch at the lateral end of the superior border transmits the suprascapular nerve, which may become entrapped here, and demarcates the bases of the coracoid process and ‘neck’ of the scapula from the remainder of the bone. The coracoid process projects mainly anteriorly in the anatomical position, and gives attachment to three muscles (coracobrachialis, short head of biceps and pectoralis minor) and three ligaments (coracoclavicular, coracoacromial and coracohumeral). The thick, cylindrical neck, to which the suprascapular nerve and vessels are very closely related posteriorly, bears the shallow articular surface for the humerus, the glenoid cavity. A tubercle immediately above the glenoid gives attachment to the long head of biceps, while the long head of triceps attaches in the corresponding position below. The spine of the scapula projects from its posterior surface, and ends laterally in the flattened acromion process. The spine is a lever for muscle attachment, with trapezius superiorly and deltoid inferiorly. The posterior surface of the body gives origin to the supraspinatus and infraspinatus muscles, both of which are tightly bound down by thick fascia. Subscapularis fills the concavity of the body anteriorly, ‘padding’ the bone against the chest wall and its covering muscles. Serratus anterior, ‘slinging’ the scapula from the upper eight ribs, attaches to the inner aspect of the medial border of the scapula. The edge of the medial border gives attachment to the rhomboids and levator scapulae, which together with trapezius connect the pectoral girdle, and thus the upper limb, with the axial skeleton. Intra-articular fractures involving the glenoid may require internal fixation, but fractures of the body of the scapula displace little and unite well, owing to the close coverage, firm fascial binding to bone, and excellent blood supply of the overlying muscles.
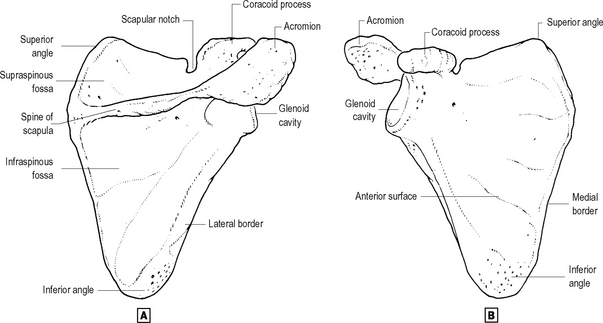
 posterior view.
posterior view.  anterior view.
anterior view.Arthrology
The ‘shoulder joint’ is not the ‘shoulder’ joint. When a patient is asked to move the shoulder, or complains of pain in the shoulder, they are not concerned only with the glenohumeral joint. The shoulder complex – the mechanism by which the upper limb is positioned and fixed in space – includes all the joints of the pectoral girdle as well as the important ligaments and muscles which link the limb to the trunk. The pectoral girdle as a whole is very mobile, movements of the ‘shoulder complex’ conferring wide versatility of function on the upper limb. The limb skeleton is linked to the trunk by a chain of synovial joints: this comprises the glenohumeral joint (see p. 355), the acromioclavicular joint and the sternoclavicular joint. The pectoral girdle is ‘slung from’ the trunk and the vertebral axis by the numerous muscles attaching to scapula and clavicle and mentioned above, while the limb is in turn connected to the girdle by a further series of muscles attaching mainly to the scapula but including pectoralis major and deltoid, attached also to the clavicle. Both pectoralis major and a further large, ‘migrant’ limb muscle, latissimus dorsi, attach the limb directly to the trunk. Note that both these muscles can act as accessory muscles of respiration if their attachment to the limb is fixed.
Neither the sternoclavicular nor the acromioclavicular joint has inherent bony stability, though both contain fibrous intra-articular discs which aid congruity. Both these synovial joints rely on their capsules and on strong, closely adjacent ligaments to provide the stability necessary for effective action of the limb (Fig. 12.8).
In the sternoclavicular joint:
In the acromioclavicular joint:
Transitional zones
Area enclosed by the pectoral girdle(s) (Fig. 12.7)
The thoracic spine, the axioscapular muscles (rhomboids and trapezii), the scapulae, clavicles and manubrium sternae form a ‘ring’ which may be considered as the complete pectoral ‘girdle’. This outer ring encompasses an inner ring made by the first ribs and their cartilages, the thoracic inlet. Thus in addition to including the major anatomical transitional zone between the neck and the thorax, this outer ring also encloses structures on their way between the root of the neck and the upper limbs (brachial plexus, subclavian vessels). There are also several surgically important named fasciae related and attached to the girdle(s) and to the boundaries of these inner and outer rings.
Between girdle and limb – the axilla
In addition to the plexus and vessels, the axilla contains fat, lymph nodes (see p. 482), the tail of the breast, the proximal parts of coracobrachialis and biceps brachii, and the intercostobrachial nerve. The key structures in the surgical anatomy of the axilla are the axillary vein (more vulnerable than the artery, and more closely related to the lymph nodes), pectoralis minor, the infraclavicular brachial plexus and the various fasciae.
Nerves
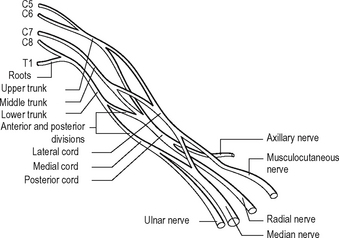
Branches of the brachial plexus
Cords
The brachial plexus can be pre-fixed or post-fixed on rare occasions. A pre-fixed plexus has a contribution from C4; a post-fixed from T2.
Clinical points
Lesions of the lower roots (C8,T1) produce the less common Klumpke’s palsy.
UPPER LIMB
Osteology
Humerus (Fig. 12.11)
The proximal expanded end of the humerus has three major components: the head and the two tuberosities or tubercles. The head, covered with articular cartilage, lies immediately above the anatomical neck. The greater tuberosity, less obviously the larger of the two when viewed from the front, lies lateral and posterior to the head. The lesser tuberosity lies anteriorly, and is separated from the greater by the bicipital (intertubercular) groove in which runs the tendon of the long head of biceps. The tendon lies in a synovial sheath which is continuous with the synovium of the glenohumeral joint. The proximal metaphysis of the humerus, below the head and tuberosities, is commonly fractured and is known as the ‘surgical neck’ of the bone. The tuberosities are processes for the attachment of the muscles of the rotator cuff, a very important group of four short muscles whose function as stabilisers of the shoulder joint overrides their function as motors of the limb. All four pass across the joint from scapula to humerus; only one lies anteriorly, and is thus a medial rotator of the humerus. This is subscapularis, attaching to the lesser tuberosity and reinforcing the anterior aspect of the joint. The other three attach to the greater tuberosity: supraspinatus, an abductor, lies above the joint and reinforces the superior capsule, while infraspinatus and teres minor lie posteriorly, are external rotators, and reinforce the posterior part of the capsule. Three other muscles pass anterior to the vertical axis of the shoulder and are thus medial rotators; all attach to the anterior aspect of the upper shaft in the region of the bicipital groove. Pectoralis major crosses the groove to its lateral lip, latissimus dorsi attaches in the floor of the groove, and teres major to its medial lip. The lineof attachment of the joint capsule excludes the tuberosities but includes the medial metaphysis, crossing the growth plate and making its medial end intracapsular. The axillary nerve lies just medial to this point, just below the capsule. The circumflex humeral arteries lie in close circumferential relationship with the bone at the surgical neck. The first important feature of the shaft of the humerus is the spiral groove, in which the radial nerve and the profunda brachii vessels run between the lateral and medial heads of triceps and in direct contact with bone. The other main feature is the tubercle for the attachment of the deltoid muscle, almost half way down the lateral border of the bone. The triangular shape of deltoid gives it its name: its broad base lies proximally on the pectoral girdle, so that it has fibres running anteriorly, superiorly and posteriorly to the shoulder joint. It can thus flex and extend the joint, in addition to its main function as an abductor. The motor supply of deltoid is the axillary nerve (C5,6), vulnerable just below the joint capsule at the surgical neck. The distal half of the front of the shaft is covered anteriorly by the attachment of brachialis and posteriorly by that of the medial head of triceps. Coracobrachialis attaches medially opposite the deltoid attachment.
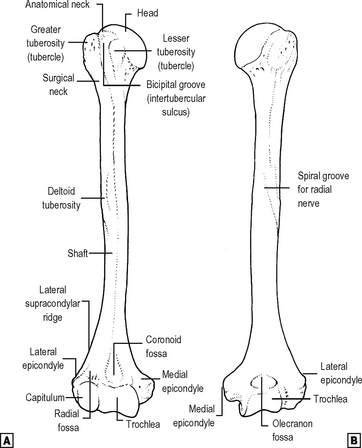
 anterior view.
anterior view.  posterior view.
posterior view.Radius (Fig. 12.12)
The radius has two expanded ends of which the distal is by far the larger. The cylindrical proximal end or head forms part both of the elbow joint and of the superior radioulnar joint, its articular surfaces for both these joints being in continuity. Immediately distal to the circumferential radioulnar articular surface of the head is the narrower neck of the bone. The annular ligament runs around this circumferential articular surface of the head, not around the neck. The main muscles attaching to the proximal radius are the biceps medially, to the bicipital tuberosity, and the supinator laterally, wrapping around the neck and proximal shaft.
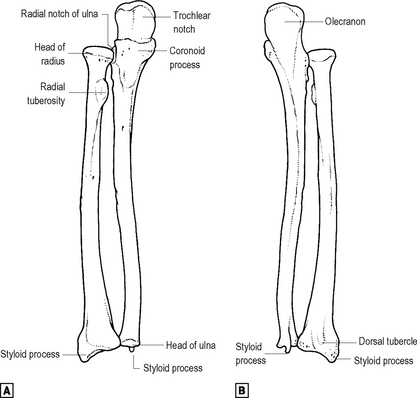
 anterior view.
anterior view.  posterior view.
posterior view.Wrist
The carpal bones (Fig. 12.13) are arranged in two rows. The proximal row, from radial (lateral) to ulnar (medial) side, comprises scaphoid, lunate and triquetral, with the pisiform, a sesamoid bone in the tendon of flexor carpi ulnaris, located anteromedially. Some features of the scaphoid, the most commonly fractured carpal bone, should be recognised, in particular the waist of the bone and the way in which the blood supply enters mainly from the distal end, making avascular necrosis of the proximal end likely after a displaced waist fracture. The distal row, again from radial to ulnar, is made up of the trapezium, trapezoid, capitate and hamate bones. The carpal bones form a shallow arch, convex dorsally. The height of this arch is increased by ventral (palmar) bony processes, the hook of the hamate, the ridge of the trapezium and the tubercle of the scaphoid, and by the pisiform medially. These ‘pillars’ of the arch give attachment to the flexor retinaculum so that the concavity of the arch forms the carpal tunnel.
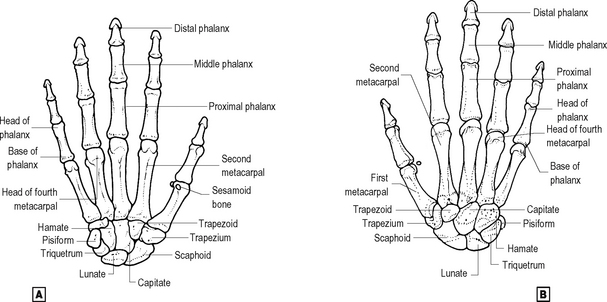
 anterior view.
anterior view.  posterior view.
posterior view.Arthrology
Glenohumeral joint
This is a ball-and-socket joint, the relative shape and size of whose articular surfaces make it totally reliant on soft tissue structures for static and dynamic stability (Fig. 12.14). The capsule is attached around the margin of the glenoid cavity of the scapula, extending onto the base of the coracoid superiorly to include the biceps attachment. The glenoid labrum, deepening the concavity of the glenoid fossa, is entirely intracapsular. On the humerus, the capsule is attached around the anatomical neck except where it passes onto the medial metaphysis inferiorly. The latter attachment brings the inferior capsule into close relation with the axillary nerve, rendering the nerve vulnerable in anteroinferior dislocations. It also means that a metaphyseal osteomyelitic lesion of the proximal humerus may be intracapsular, leading to the possibility of septic arthritis as a sequel. The capsule is reinforced by the tendons of the rotator cuff muscles, which blend with it everywhere except inferiorly, and additionally by the coracohumeral ligament superiorly. There are also variable thickenings in the capsule anteriorly: these are the glenohumeral ligaments (GHL), whose detailed anatomy is of importance to the shoulder arthroscopist. The capsule is lax inferiorly, allowing wide abduction of the joint. The tendon of the long head of biceps passes through the joint over the head of the humerus, within its synovial sheath: it is intracapsular but extrasynovial (cf. the cruciate ligaments of the knee) and is a major arthroscopic landmark. The synovial sheath extends distally beneath the transverse ligament of the humerus into the bicipital groove. The two major bursae associated with the joint are the subacromial/subdeltoid bursa superiorly and the subscapular bursa anteriorly. The former lies between the ‘layers’ of the abductor mechanism of the shoulder (the acromion, with the attached deltoid muscle and coracoacromial ligament, and supraspinatus attaching to the greater tuberosity). It does not normally communicate with the synovial cavity of the glenohumeral joint. The latter is an extension of the glenohumeral synovium passing between the glenohumeral ligaments and deep to subscapularis muscle. The muscles crossing the glenohumeral joint and their actions upon it are summarised below. The joint is innervated by the nerves which supply those muscles, mainly by the axillary and suprascapular. The suprascapular artery, from the subclavian, and the subscapular and circumflex humeral arteries, from the axillary, are the main participants in the anastomosis around the scapula and the head of the humerus. The glenohumeral capsule is said to be at its tightest when the joint is abducted and externally rotated, though this position is often that which produces anteroinferior dislocation. The joint may be aspirated or injected anteriorly or posteriorly, the posterior subacromial approach being somewhat easier and also being that usually used for arthroscopy. Diagnostic arthrography of the shoulder has been largely superseded by MRI. The common approach for open shoulder surgery is anterior, passing between deltoid and pectoralis major. The muscles attaching to the coracoid are displaced medially, protecting axillary neurovascular structures, and the capsule is entered after dividing subscapularis.
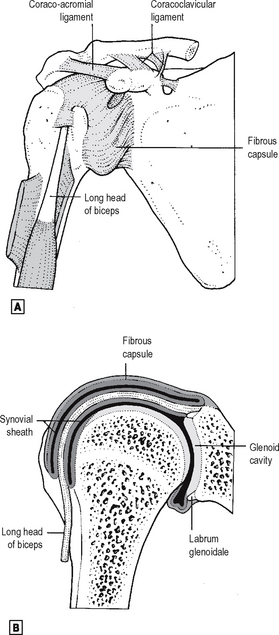
 anterior view.
anterior view. in section.
in section.Elbow joint
This is a modified hinge joint between the humerus and the forearm bones (Fig. 12.15). The main part of the joint is the humero-ulnar, between the trochlea of the humerus and the trochlear notch of the ulna. The trochlear diameter is greater medially than laterally, thus creating the valgus ‘carrying angle’ of the elbow. The trochlear articular surface of the humerus is continuous laterally with that of the rounded capitulum, which is confined to the anterior aspect of the bone and which articulates with the concavity of the head of the radius. The trochlear articular surface of the ulna continues laterally and distally over the radial notch, which articulates with the circumferential part of the radial articular surface. The superior radio-ulnar joint is thus continuous with the elbow joint, sharing a synovial ‘cavity’. The humeral attachment of the capsule of the elbow joint leaves the articular margins of trochlea and capitulum anteriorly and posteriorly to include the coronoid, radial and olecranon fossae. The capsule is thin anteriorly and posteriorly but thicker medially and laterally where reinforced by the collateral ligaments. It attaches distally to the articular margins of the trochlear notch, then passes onto the superior border of the annular ligament, with which it becomes continuous. The collateral ligaments of the elbow differ in shape: the radial is fan-shaped, radiating onto the annular ligament from the lateral humeral epicondyle, while the ulnar has three bands constituting a triangle between the medial epicondyle and the lateral sides of the coronoid and olecranon processes (Fig. 12.15). The synovium of the elbow, lining the capsule, is continuous with that of the superior radio-ulnar joint, and extends distally a little way below the annular ligament. The synovium is separated from the capsule anteriorly and posteriorly by fat pads, whose displacement can be used in the radiological diagnosis of small effusions or haemarthroses of the elbow. Muscles crossing, and thus acting upon, the joint are summarised below. The nerves to the elbow derive, by Hilton’s Law, from all three major nerves of the limb. The arterial anastomosis around the elbow is made up of branches from the brachial, radial and ulnar vessels. The elbow is maximally stable in full extension, when the anterior capsule is tense. A swollen elbow is best aspirated from the lateral side, where no important neurovascular structures cross the joint. A posterior approach, lateral to the main body of the triceps tendon, may also be used. Open surgical approaches are mainly lateral or posterior, depending upon the size of exposure required.
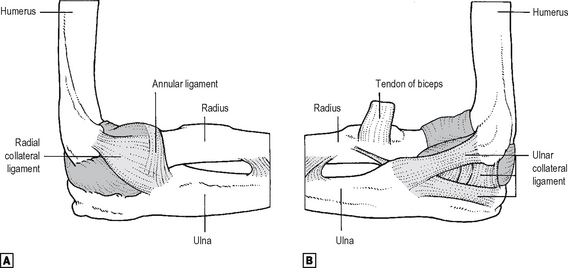
 lateral view.
lateral view.  medial view.
medial view.Radio-ulnar joints
The shafts of radius and ulna are strongly linked in all positions of the forearm by the interosseous membrane, whose fibres pass distally and medially, transmitting force from radius to ulna. The proximal ends of the bones articulate at the superior radio-ulnar joint, described above with the elbow, with which it is structurally continuous. The distal ends articulate at the inferior radio-ulnar joint, between the cylindrical ulna and the concave ulnar notch of the radius. The capsule of this joint is weak, but the joint is strengthened by a triangular intra-articular fibrocartilage passing between the ulnar styloid and the distal margin of the ulnar notch of the radius. If intact, this fibrocartilage separates the synovial lining of the inferior radio-ulnar joint from that of the wrist joint. The radio-ulnar joints move together during forearm rotation into pronation and supination, the axis passing from the centre of the radial head to theulnar styloid.
 anterior view.
anterior view.  posterior view.
posterior view.Between the joints: fasciae and compartments; nerves
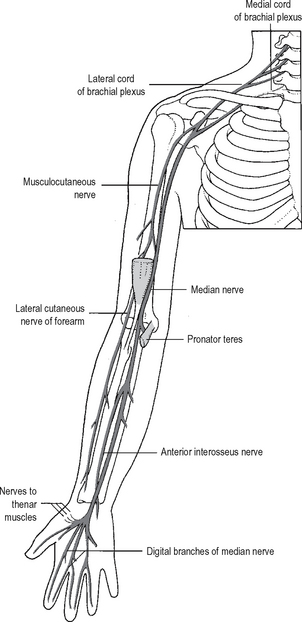
Fig. 12.16 The course and distribution of the median and musculocutaneous nerves.
Source: Rogers op. cit.
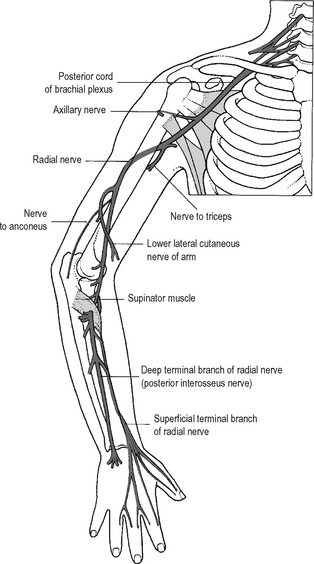
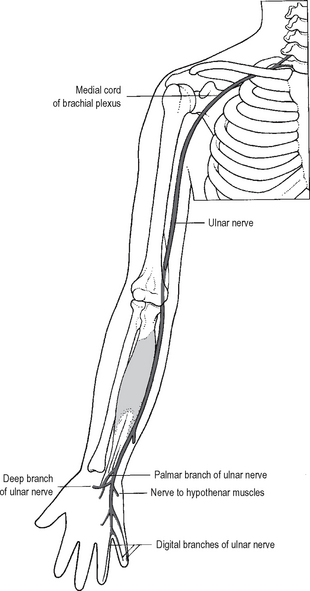
The median nerve (Fig. 12.16) and the brachial artery remain in the flexor compartment. The ulnar nerve (Fig. 12.18) lies in the flexor compartment proximally but passes into the extensor compartment before crossing the elbow. The radial nerve does exactly the reverse, lying anterolaterally at the elbow.
Transitional zones
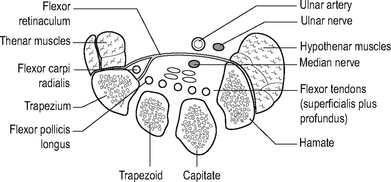
Carpal tunnel
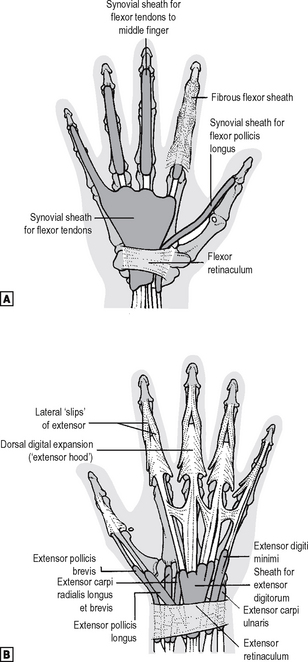
The flexor retinaculum forms the roof of a tunnel, the floor and walls of which are made up by the concavity of the carpal bones (Fig. 12.19). Within this tunnel are the tendons of flexor digitorum superficialis, flexor digitorum profundus, flexor pollicis longus and flexor carpi radialis (the latter tendon is in its own separate osseofascial compartment. The most important structure to pass through the tunnel is the median nerve. Any lesion diminishing the size of the tunnel may result in compression of the median nerve (carpal tunnel syndrome). The superficial palmar branch of the nerve is given off proximal to the flexor retinaculum and, therefore, there is no sensory impairment on the lateral side of the palm if the nerve is compressed in the carpal tunnel.
Anatomical snuff box
The anatomical snuff box is formed as follows:
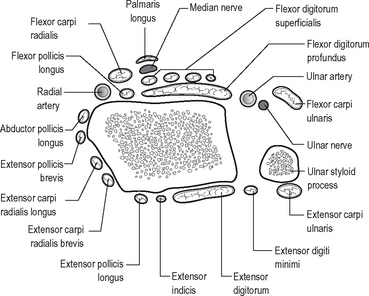
Relationships around the wrist are important as this is a commonly injured region. The structures around the wrist joint are shown in Fig. 12.20.
Hand
On the palmar side note first the anatomy of the carpal tunnel. Its boundaries have been described in the osteology section above. Note its surface marking: the proximal edge of the flexor retinaculum underlies the distal wrist crease, so that the tunnel lies effectively in the palm. Running through the tunnel are the eight long finger flexor tendons, lying in a common synovial sheath which is incomplete radially. This common sheath extends distally into the palm (Fig. 12.21). The superficialis tendons run in two pairs, those to the middle and ring fingers lying nearer the retinaculum. The profundus tendons run more dorsally in a single row of four, that to the index being separate from the medial three. The tendon of flexor pollicis longus (FPL) runs through the tunnel radial to the finger flexors, in its own synovial sheath. The median nerve lies just beneath the retinaculum, radial to its midpoint. Also related to the retinaculum are the ulnar nerve and vessels and the tendon of flexor carpi radialis (FCR). The ulnar nerve and vessels run superficial to theretinaculum just radial to the pisiform, while the FCR tendon lies in its own osteofascial compartment inthe tunnel superficial and radial to the FPL. The retinaculum is also crossed superficially near its midpoint by the palmar cutaneous branch of the median nerve. The muscles of the thenar and hypothenar eminences attach proximally to the radial and ulnar sides of the palmar surface of the retinaculum. The thenar eminence contains abductor pollicis brevis superficially, with flexor pollicis brevis (FPB) and opponens pollicis lying more deeply. All are usually supplied by the motor branch of the median nerve, given off as that nerve leaves the carpal tunnel, though FPB and opponens may be supplied by the ulnar. The hypothenar eminence contains a similarly named trio of muscles:abductor, flexor and opponens digiti minimi. All are supplied by the ulnar nerve. In both eminences, all except the opponens muscles attach distally to the base of the proximal phalanx. The opponentes attach to their respective metacarpals. The central part of the palmar surface of the flexor retinaculum is overlaid by the proximal ‘apex’ of the triangular palmar fascia: the tendon of palmaris longus attaches to this apex. Thin expansions from the radial and ulnar sides of the palmar fascia cover the thenar and hypothenar muscles. A septum from the dorsal surface of the fascia passes to the shaft of the third (middle) metacarpal, dividing the potential space deep (dorsal) to the common flexor synovial sheath into two compartments. These are the palmar spaces: the midpalmar medially and the thenar laterally (see p. 401 and Fig. 12.42). The triangular adductor pollicis muscle lies deep to the thenar space, its base attaching, like the fascial septum, to the third metacarpal shaft, and its apex to the proximal phalanx of the thumb. Attached to the radial sides of the deep finger flexor tendons as they lie in the common sheath in the palm are the lumbrical muscles, each running distally to attach to a digital extensor expansion. Theselittle muscles control the relative tension in the long finger flexors, and help the interossei in flexing the MCP joints. Deeper still lie the interosseous muscles, attached as shown in Fig. 12.22. From the level of the MCP joints distally into the digits, the flexors for each digit share a fibrous sheath which surrounds the synovial sheath. This fibrous sheath is subdivided into thick bands or ‘pulleys’ opposite the phalangeal shafts, and thinner, flexible areas opposite the joints. On the dorsal side the only muscle belly easily palpable is that of the first dorsal interosseous. This, with adductor pollicis, forms the bulk of the web between thumb and index. As both muscles are innervated by the ulnar nerve, weakness and wasting here are important diagnostic signs. The tendons of the digital extensorspass in their own synovial sheaths beneath the extensor retinaculum on the dorsum of the wrist (Fig. 12.21). Note that the extensors and long abductor of the thumb bound the ‘anatomical snuff-box’, in whose baselies the scaphoid bone crossed by the radial artery. Each finger extensor inserts into an arrowhead-shaped fibrous ‘hood’, which wraps around the dorsum of the finger (Fig. 12.21). The thumb extensors insert more directly into bone. The intrinsic muscles of the digits (lumbricals and interossei) insert into the digitalextensor hoods from the flexor side (Fig. 12.22), so that as they contract together they tighten the hood around the proximal phalanx and thus flex the MCP joint. The main body of the extensor tendon attaches to the base of the middle phalanx, while its lateral extensions come together to attach to the distal phalanx (Fig. 12.22).
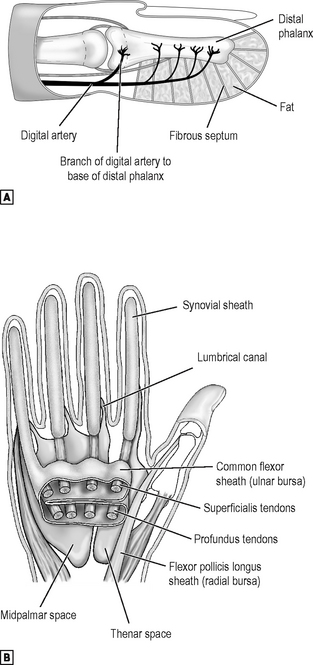
 a diagram of the pulp space.
a diagram of the pulp space.  the left palmar spaces and synovial sheaths. Infection inthe thenar or midpalmar spaces easily breaks through into the lumbrical canals (connective tissue sheath of the lumbrical muscles), so the canals are shown in continuity with the spaces. A segment of the ulnar bursa has been removed to indicate the disposition of the finger tendons embraced by it.
the left palmar spaces and synovial sheaths. Infection inthe thenar or midpalmar spaces easily breaks through into the lumbrical canals (connective tissue sheath of the lumbrical muscles), so the canals are shown in continuity with the spaces. A segment of the ulnar bursa has been removed to indicate the disposition of the finger tendons embraced by it.Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


