Part 1 Introduction
Terminology
It is essential for students beginning their study of anatomy to become familiar with an internationally accepted vocabulary, allowing communication and understanding between all members of the medical and paramedical professions throughout the world. Perhaps the single, most important descriptive feature of this vocabulary is the adoption of an unequivocal position of the human body. This is known as the anatomical position. It is described as follows: the body is standing erect and facing forwards; the legs are together with the feet parallel so that the toes point forwards; the arms hang loosely by the sides with the palm of the hand facing forwards so that the thumb is lateral (Fig. 1.1). All positional terminology is used with reference to this position, irrespective of the actual position of the body when performing an activity.
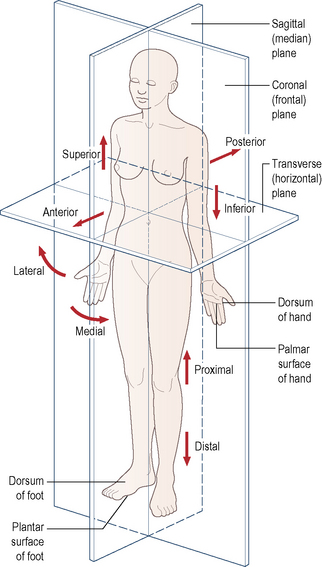
To facilitate the understanding of the relation of structures one to another and the movement of one segment with respect to another, imaginary reference planes pass through the body in such a way that they are mutually perpendicular to each other (Fig. 1.1). Passing through the body from front to back and dividing it into two symmetrical right and left halves is the sagittal (median) plane. Any plane parallel to this is also known as a parasagittal (paramedian) plane.
Terms used in describing movement
Supination and pronation are terms used in conjunction with the movements of the forearm and foot.
Inversion and eversion are terms used to describe composite movements of the foot.
Nervous system
Introduction
The nervous system consists of highly specialized cells designed to transmit information rapidly between various parts of the body. Topographically it can be divided into two major parts: the central nervous system (CNS) and the peripheral nervous system (PNS). The brain and spinal cord constitute the CNS, which lies within the skull and vertebral canal (see p. 572 and p. 489), while nerves in the PNS connect the CNS with all other parts of the body (see p. 7).
Cellular structure
The basic cellular unit of the nervous system is the nerve cell or neuron, which differ in size and shape according to their function and location within the nervous system. All neurons have three characteristic components: a cell body, an axon, and dendrites (Fig. 1.2A).
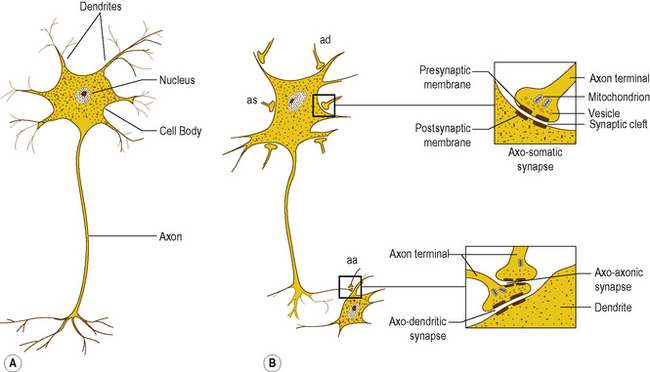
Interneural connections
Synapses are formed by the close approximation of a small discrete area of the cell membrane of one neuron to a reciprocal area of the membrane of a second neuron (Fig. 1.2B). The apposed membranes are specially modified and are separated by a narrow gap about 0.02 μm in width, called the synaptic cleft. Across the synapse one cell communicates with the next, with communication being unidirectional. The membrane of the cell transmitting the information is the presynaptic membrane, and that of the cell receiving the information the postsynaptic membrane.
Myelination
In the PNS, myelination is by cells called Schwann cells: it is achieved by a Schwann cell curling an extension of its cell membrane around the shaft of the axon in a spiral manner (Fig. 1.3A). As the Schwann cell extension wraps around the axon, the cytoplasm in it is squeezed out until only a double layer of the cell membrane of the Schwann cell remains spiralling around the axon (Fig. 1.3A). In this way the myelin sheath is formed by the lipid and protein of the cell membrane of the Schwann cell that surrounds the axon. Peripheral to the myelin sheath, the axon is surrounded by the cytoplasm of the Schwann cell, with the outermost cell membrane of the Schwann cell acting as a second membrane to the axon; this forms the neurolemma (Fig. 1.3A).
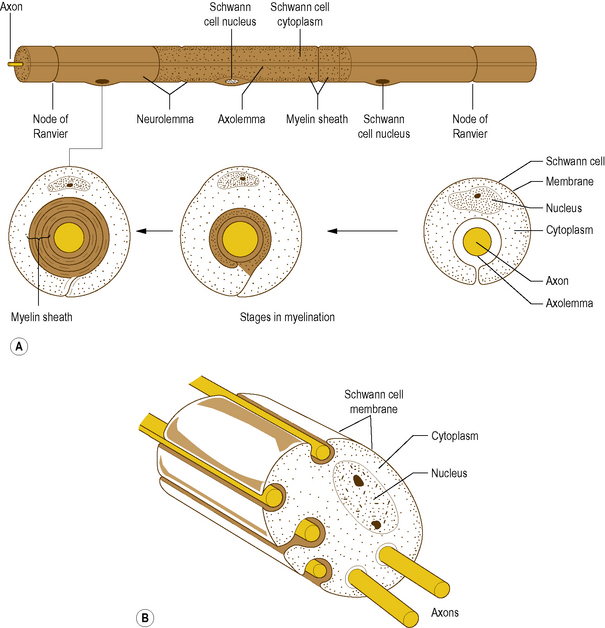
Along its length, a given axon is ensheathed in series by a large number of Schwann cells, but each Schwann cell is related only to a single axon (Fig. 1.3A). Junction points where the myelin sheath of one Schwann cell ends and the sheath of the next begins are known as Nodes of Ranvier. An axon together with the myelin sheath and the Schwann cells that surround it is referred to as a nerve fibre.
While many axons in the PNS are myelinated a large number remain unmyelinated. Instead of being enveloped by a tightly spiralling sheath of myelin, they run embedded in invaginations of Schwann cell membranes, so that a single Schwann cell may envelop several axons (Fig. 1.3B). Unmyelinated axons are afforded less physical protection than myelinated ones, but the foremost difference between them is that unmyelinated axons conduct impulses at much slower velocities.
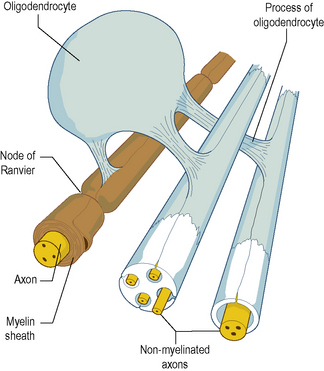
In the CNS, myelination is performed by specialized cells called oligodendrocytes. The process is basically similar to that in the PNS, except that a given oligodendrocyte is usually involved in the simultaneous myelination of several separate axons (Fig. 1.4). Unmyelinated axons in the CNS run embedded in cytoplasmic extensions, or processes, of oligodendrocytes.
Structure of a peripheral nerve
Individual myelinated axons are surrounded by a tubular sheath of fibrous tissue, the endoneurium, whereas clusters of axons are held together by a larger fibrous sheath, the perineurium (Fig. 1.5). Unmyelinated axons are not enclosed by endoneurium but run in isolated bundles parallel to myelinated axons and are enclosed with them in the perineurial sheath.
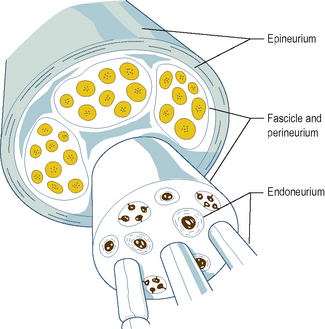
A bundle of axons enclosed within a single perineurial sheath is referred to as a nerve fascicle (Fig. 1.5). Axons within a fascicle largely remain within that same fascicle throughout the course of the nerve until their peripheral distribution. However, along the course of a peripheral nerve axons may at times leave one fascicle to enter and continue within an adjacent fascicle.
The fascicles within a peripheral nerve are bound together by an external sheath of fibrous tissue, the epineurium (Fig. 1.5), which forms the external surface of the macroscopic nerve. As a peripheral nerve passes through the tissues of the body, it gives off branches composed of one or more fascicles of the parent nerve, which leave it to reach their particular destination. Along the course of a nerve, this process is repeated until all the fascicles and axons in the nerve have been distributed to their target tissues.
The peripheral nervous system
The nerves that supply the viscera, such as the heart, lungs and digestive tract, are referred to as visceral nerves. However, because the nervous functions concerning viscera are largely automatic and subconscious, that part of the nervous system that innervates viscera is referred to as the autonomic nervous system (ANS). It has components in both the central and PNSs: it is described in detail on page 500.
Constituents of peripheral nerves
Axons may also be classified according to their conduction velocities, which in turn are proportional to their sizes (Fig. 1.6). If a peripheral nerve is stimulated electrically and recordings of the evoked activity are taken some distance along the nerve, a wave of electrical activity can be recorded. The wave is generated by the summation of the electrical activity in each of the axons in the nerve, its size being proportional to the number of axons in the nerve, while its shape reflects the type of axons present.
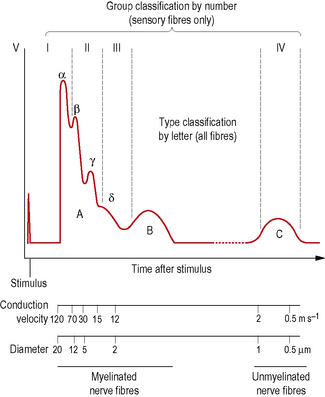
The waveform obtained from a nerve containing every known type of axon is represented in Fig. 1.6. It has three principal peaks, named the A, B and C waves. The A wave occurs very soon after the triggering stimulus, as it is produced by rapidly conducting axons with velocities in the range 12–120 ms−l: these are classified as A fibres. The A wave is broken down into several secondary peaks called Aα, Aβ, Aγ and Aδ waves, each being produced by a particular subgroup of rapidly conducting axons.
Another system of axon classification relates specifically to sensory fibres. In this system, sensory fibres are classified according to their conduction velocities into groups I, II, III and IV. These groups have the same conduction velocities as type Aα, Aβ, Aδ and C fibres respectively, as shown in Fig. 1.6, but do not include the Aα and Aβ motor fibres. The fibres of groups III and IV largely mediate pain and temperature sensations. Group II constitutes fibres that mediate pressure and touch, and fibres that form spray endings in muscle spindles (p. 15). Group I is divided into groups Ia and Ib. Group Ia fibres are slightly larger and innervate muscle spindles, while group Ib fibres innervate Golgi tendon organs (p. 15).
Nerve endings
The terminals of axons in peripheral nerves have unique structures depending on their function. Motor axons have terminals designed to deliver a stimulus to muscle cells (see p. 16), while sensory axons have terminals designed to detect particular types of stimuli. These terminals are called receptors, and a diversity of morphological types can be found (see p. 26).
Nervous system
Components of the musculoskeletal system
Skeletal tissue
Bone
The internal architecture of bone reveals a system of struts and plates (trabeculae) running in many directions (Fig. 1.7), which are organized to resist compressive, tensile and shearing stresses. Surrounding these trabecular systems, which tend to be found at the ends of long bones, is a thin layer of condensed or compact bone (Fig. 1.7). The network of the trabeculae, because of its appearance, is known as cancellous or spongy bone. In the shaft of a long bone there is an outer, relatively thick ring of compact bone surrounding a cavity, which in life contains bone marrow.
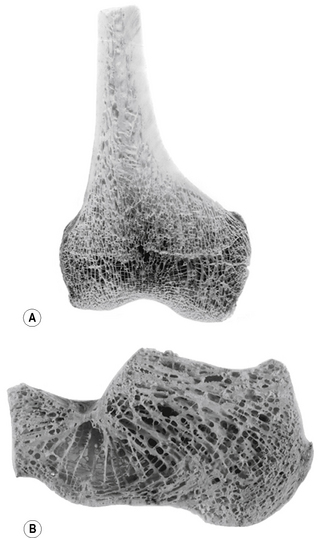
For descriptive purposes bones can be classified according to their shape:
Both irregular and short bones consist of a thin layer of compact bone surrounding cancellous bone.



