(1)
Faculty of Life Sciences, Bar-Ilan University, Ramat-Gan, Israel
Keywords
Mitochondrial functionNADH fluorescenceIn vivo monitoringOxygen balanceMultiparametric monitoring1.1 The Mitochondrion
The understanding of mitochondrial function has been a challenge for various investigators since its discovery more than 150 years ago. In 1981, Ernster and Schatz [1] reviewed the history of mitochondrial structure and function studies. There is no real single answer regarding who discovered mitochondria. The process of discovery and identification was a gradual one that has spanned the past 150 years.
The four scientists who were involved in the discovery of the mitochondria are presented in Fig. 1.1. In 1856, the Swiss anatomist and histologist Albert von Kölliker [2] described what he called “granules” in the sarcoplasm of striated muscle. Retzius in 1890 called the granules “sarcosomes,” which later were identified as the mitochondria [3].

Fig. 1.1
The four scientists who were involved in the discovery of the mitochondria
The discovery of mitochondria in general came in 1890 when Richard Altmann, a cytologist, identified the organelles and dubbed them “bioblasts” [4]. Carl Benda, in 1898, coined the term mitochondria from Greek thread, ‘mitos,’ and granule, ‘chondros’ [5].
As seen in Fig. 1.2, disturbances of mitochondrial function lead to disruption of cellular activities, inducing various human diseases [6] such as neurodegenerative processes, which are related to apoptosis [7–9]. The aging process has also been related to mitochondrial dysfunction [10, 11]. Moreover, mitochondrial dysfunction is involved in the pathologies of the nervous system, such as traumatic brain injury [12, 13], spinal cord injury [14], and stroke [15]. The implication of mitochondrial dysfunction in cardiovascular diseases was recently reviewed [16]. The relationship between mitochondrial activity and prolonged endotoxemia in the liver, skeletal muscle, and kidney has been reported [17, 18], as well as its role in sepsis [12, 19]. Metabolic disorders, including visceral obesity, diabetes mellitus, dyslipidemia, nonalcoholic fatty liver disease, and hypertension, are also connected with mitochondrial dysfunction [20–22]. The neurotoxic effects of drug abuse are often associated with oxidative stress and mitochondrial dysfunction, among other mechanisms [23, 24].
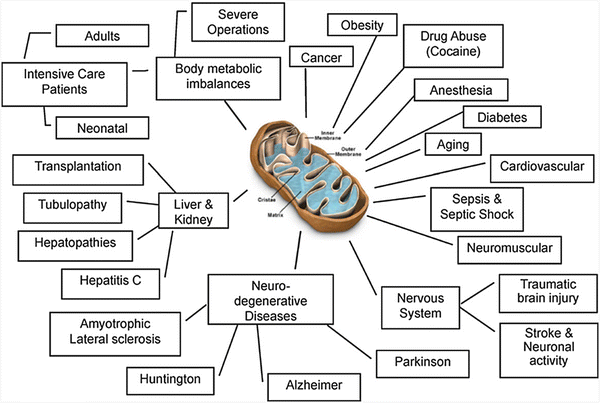
Fig. 1.2
Involvement of mitochondrial dysfunction in various pathophysiological conditions. Mitochondrial dysfunction was described as a critical step in the development of many states of disease in patients. (© Reprinted with permission from Elsevier [48])
The mitochondria, as an energy production system, are involved in tumor cell pathogenesis, initially described by Warburg 80 years ago [25] and later followed by many studies [8, 26–29]. The role of NADH in cellular function and cell death as well as in brain functions, diseases, and aging was reviewed in 2006 [30, 31]. The connection between mitochondria and neuronal activity was described in detail in 2007 [32]. In 2013, Edeas and Weissig published a paper claiming that the future of medicine will come through mitochondria [33].
The role of NADH in regulating cellular metabolism was described by Kaplan [34].
A general, short overview on the connection between cancer cells and cellular energy metabolism is presented here. The interrelation between the two components of cellular energy metabolism, namely, glycolysis and mitochondrial oxidative phosphorylation, was first studied by Louis Pasteur in 1857 [35]. Pasteur found that when oxygen is provided to cells metabolizing anaerobically, O2 consumption increases (the Pasteur effect), and utilization of glucose and production of lactic acid declines. The “aerobic glycolysis” in cancer cells, described by Warburg in the 1920s, was contrary to the normal Pasteur effect. Warburg claimed that in cancer cells glycolysis is the main source of ATP even when oxygen supply is normal because mitochondrial function is inhibited. The Crabtree effect, described about the same time [36, 37], was tested in the mid-1960s by Bickis and Henderson [38] and Bickis et al. [39] in sliced tissues in vitro. They tried to estimate tumor malignancy from metabolic measurements as well as the sensitivity of tumors to anticancer agents. Although mitochondrial energy metabolism in cancer cells was the main subject of Warburg’s study 80 years ago, the investigation of his hypothesis by real-time monitoring of mitochondrial function in vivo was relatively neglected during the past 30 years. This issue was discussed in detail by Gatenby and Gillies [40], who suggested new avenues of investigation related to critical issues of the “relationship in timing between angiogenesis switch and the glycolytic switch.” Involvement of the mitochondria and angiogenesis in hematological malignancies were discussed by various investigators [41–44].
During these past few decades, the dominant approach in research on the involvement of mitochondria in cancer and tumor development focused on signal transduction and other nonenergetic aspects, such as apoptosis and reactive oxygen species (ROS) generation. These various investigation subjects were based on Warburg’s assumption that cancer could be the result of mitochondrial defects. Brandon’s group concluded that “mitochondrial dysfunction does appear to be a factor in cancer etiology, an insight that may suggest new approaches for diagnosis and treatment” [28].The actuality of Warburg relevance to renal cancer metabolism was discussed by Simonnet et al. [45] and Godinot et al. [46]. Figure 1.3 presents six main subjects that are relevant to mitochondrial function in cancer cells and tumors [47]. It is important to note that this classification was designed to show the complex relationship between mitochondrial activity and cancer in a simplified scheme. Each item presented in the figure has been deeply investigated, with a large number of experimental and clinical studies published during the last few decades. The multiple interconnections between each of the presented items made it almost impossible to draw lines between them. The details of the six subjects appeared in our published paper [47].
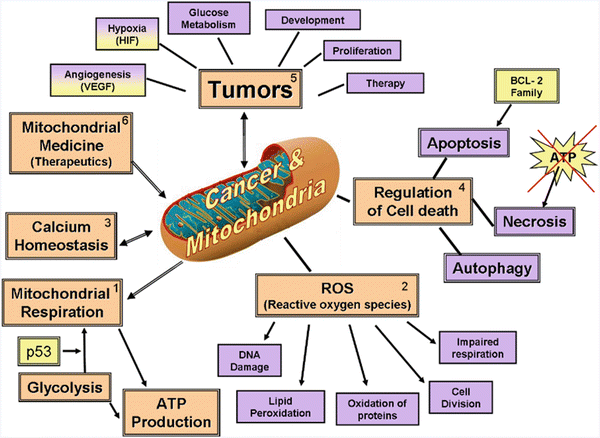
Fig. 1.3
Central role of the mitochondrion in various processes involved in the pathology of cancer cells and tumors. Six issues marked as 1–6 are discussed in the published paper. (© Reprinted with permission from Elsevier [47])
Therefore, the possibility of monitoring mitochondrial function in vivo, in experimental animals and in patients, is of great importance and could contribute significantly to the understanding of various pathological processes.
Most of the information on mitochondrial function has been accumulated from in vitro studies. A relatively small portion of published papers involved the monitoring of mitochondrial function in vivo and in real time. During the past 40 years, we have published more than 150 papers in this very significant area, including the largest number of studies using NADH redox state monitoring in patients as well. In this book, I survey the various aspects of in vivo real-time monitoring of NADH in experimental animals as well as in patients. To demonstrate diverse scientific implications of in vivo monitoring, we also present selected published in vitro data.
Most of the figures presented in the book were published previously in our papers since 1972.
At the beginning of each chapter or new subject, one or two representative figures published by other investigator(s) are presented, followed by an explanation of the figure.
1.2 Collaboration of Avraham Mayevsky (A.M.) with Britton Chance (B.C.)
My first meeting with Prof. Britton Chance was in Israel during his attendance at the Biophysical Society meeting in 1971. He came to visit my advisor regarding my Ph.D. thesis, Prof. David Samuel, at the Isotope Department of the Weizmann Institute of Science, Israel. He saw the brain in vivo monitoring system for evaluation in radioactive phosphate and immediately offered me to come to the Johnson Research Foundation in Philadelphia and join his group as a postdoctoral fellow.
After my graduation from the Weizmann Institute (Rehovot, Israel) in October 1972, my family (wife and three children) came to Philadelphia and stayed there for 2 years.
After 2 years of postdoctoral activity I went back to Bar Ilan University in Israel, but our collaboration continued, and the next visit for a year was in 1980–1981. Every year between 1974 and 1988 I visited the Johnson Research Foundation for an average period of 1 month. Later on, we spent 2 years in Philadelphia during which our first attempt to monitor neurosurgical patients came through.
During my collaboration with Prof. Chance we published 33 papers together, in addition to more than 100 papers that I published with other collaborators, on NADH monitoring. Our meeting in 2007–2008 in China was, in a sense, closing a life cycle that had started in Philadelphia in 1972 and ended in the famous Britton Chance Center for Biomedical Photonics, Wuhan National Laboratory for Optoelectronics, Huazhong University of Science and Technology, Wuhan, China.
Before describing my personal anecdotes with Prof. Chance, I wish to summarize his contribution to the field of bioenergetics by monitoring mitochondrial signals using light (Fig. 1.4). He was the first to develop in detail an optical technique for the monitoring of mitochondrial signals, namely, NADH and flavoproteins in intact tissues, and later on under in vivo conditions. Brit started his activities in this field after the discoveries of Warburg and Keilin before 1950. He was the leader of modern biophotonics regarding the theoretical, experimental, and clinical application of optical monitoring of mitochondrial signals. During the first decade (1951–1962), Brit investigated the isolated mitochondria via tissues in vitro and finally under in vivo conditions. It is impossible to imagine the development of this field of bioenergetics without the foundations established by Prof. Chance. In the 1970s, he started to use the optical technology for clinical applications.

Fig. 1.4
Citation from the Old Bible regarding the creation of light
I would like to mention a few events and episodes that represent my unique personal ties with Prof. Chance (Fig. 1.5), who was my second father (scientific).

Fig. 1.5
Use of light in studying mitochondrial function was introduced by Prof. Britton Chance more than 60 years ago
1.
In October 1972 we arrived in Philadelphia with three children after an 18-h flight, and the Chance family hosted my family in their home for more than a week. This impressive welcome was very important in the establishment of my collaboration mode with Prof. Chance for more than 35 years. During this week we had an opportunity to be in daily touch with the Chance family. Before dinner, Prof. Chance (or one of the children) played the piano, which led to a very special atmosphere. Figure 1.5 shows a few pictures of Prof. Chance on various occasions.
2.




Prof. Chance was a very demanding scientist, from himself as well as from his collaborators. One day when I was looking for the nitrogen cylinder at 7 p.m. during my experiments, Brit went with the cart to his lab and brought the cylinder to my lab. This kind of behavior stimulated our activities and fruitful collaboration. One day he said, “I am waiting for your results for more than 10 years and I am pleased that you are running your studies intensively.”
Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
