30
Genetics & Gene Therapy
CHAPTER CONTENTS
INTRODUCTION
The study of viral genetics falls into two general areas: (1) mutations and their effect on replication and pathogenesis; and (2) the interaction of two genetically distinct viruses that infect the same cell. In addition, viruses serve as vectors in gene therapy and in recombinant vaccines, two areas that hold great promise for the treatment of genetic diseases and the prevention of infectious diseases.
MUTATIONS
Mutations in viral DNA and RNA occur by the same processes of base substitution, deletion, and frameshift as those described for bacteria in Chapter 4. Probably the most important practical use of mutations is in the production of vaccines containing live, attenuated virus. These attenuated mutants have lost their pathogenicity but have retained their antigenicity; therefore, they induce immunity without causing disease.
There are two other kinds of mutants of interest. The first are antigenic variants such as those that occur frequently with influenza viruses, which have an altered surface protein and are therefore no longer inhibited by a person’s preexisting antibody. The variant can thus cause disease, whereas the original strain cannot. Human immunodeficiency virus and hepatitis C virus also produce many antigenic variants. These viruses have an “error-prone” polymerase that causes the mutations. The second are drug-resistant mutants, which are insensitive to an antiviral drug because the target of the drug, usually a viral enzyme, has been modified.
Conditional lethal mutations are extremely valuable in determining the function of viral genes. These mutations function normally under permissive conditions but fail to replicate or to express the mutant gene under restrictive conditions. For example, temperature-sensitive conditional lethal mutants express their phenotype normally at a low (permissive) temperature, but at a higher (restrictive) temperature, the mutant gene product is inactive. To give a specific example, temperature-sensitive mutants of Rous sarcoma virus can transform cells to malignancy at the permissive temperature of 37°C. When the transformed cells are grown at the restrictive temperature of 41°C, their phenotype reverts to normal appearance and behavior. The malignant phenotype is regained when the permissive temperature is restored.
Note that temperature-sensitive mutants have now entered clinical practice. Temperature-sensitive mutants of influenza virus are now being used to make a vaccine, because this virus will grow in the cooler, upper airways where it causes few symptoms and induces antibodies, but it will not grow in the warmer, lower airways where it can cause pneumonia.
Some deletion mutants have the unusual property of being defective interfering particles. They are defective because they cannot replicate unless the deleted function is supplied by a “helper” virus. They also interfere with the growth of normal virus if they infect first and preempt the required cellular functions. Defective interfering particles may play a role in recovery from viral infection; they interfere with the production of progeny virus, thereby limiting the spread of the virus to other cells.
INTERACTIONS BETWEEN VIRUSES
When two genetically distinct viruses infect a cell, three different phenomena can ensue.
(1) Recombination is the exchange of genes between two chromosomes that is based on crossing over within regions of significant base sequence homology. Recombination can be readily demonstrated for viruses with double-stranded DNA as the genetic material and has been used to determine their genetic map. However, recombination by RNA viruses occurs at a very low frequency, if at all. Reassortment is the term used when viruses with segmented genomes, such as influenza virus, exchange segments. This usually results in a much higher frequency of gene exchange than does recombination. Reassortment of influenza virus RNA segments is involved in the major antigenic changes in the virus that are the basis for recurrent influenza epidemics.
(2) Complementation can occur when either one or both of the two viruses that infect the cell have a mutation that results in a nonfunctional protein (Figure 30–1). The nonmutated virus “complements” the mutated one by making a functional protein that serves for both viruses. Complementation is an important method by which a helper virus permits replication of a defective virus. One clinically important example of complementation is hepatitis B virus providing its surface antigen to hepatitis delta virus, which is defective in its ability to produce its own outer protein.
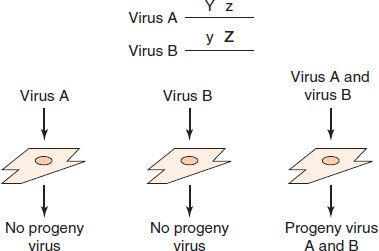
FIGURE 30–1 Complementation. If either virus A or virus B infects a cell, no virus is produced because each has a mutated gene. If both virus A and virus B infect a cell, the protein product of gene Y of virus A will complement virus B, the protein product of gene Z of virus B will complement virus A, and progeny of both virus A and virus B will be produced. Note that no recombination has occurred and that the virus A progeny will contain the mutated z gene and the virus B progeny will contain the mutant y gene. Y, Z, functional genes; y, z, mutated, nonfunctional genes.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


