4
Genetics
CHAPTER CONTENTS
INTRODUCTION
The genetic material of a typical bacterium, Escherichia coli, consists of a single circular DNA molecule with a molecular weight of about 2 × 109 and is composed of approximately 5 × 106 base pairs. This amount of genetic information can code for about 2000 proteins with an average molecular weight of 50,000. The DNA of the smallest free-living organism, the wall-less bacterium Mycoplasma, has a molecular weight of 5 × 108. The DNA of human cells contains about 3 × 109 base pairs and encodes about 100,000 proteins.
Note that bacteria are haploid; in other words, they have a single chromosome and therefore a single copy of each gene. Eukaryotic cells (such as human cells) are diploid, which means they have a pair of each chromosome and therefore have two copies of each gene. In diploid cells, one copy of a gene (allele) may be expressed as a protein (i.e., be dominant), whereas another allele may not be expressed (i.e., be recessive). In haploid cells, any gene that has mutated—and therefore is not expressed—results in a cell that has lost that trait.
MUTATIONS
A mutation is a change in the base sequence of DNA that usually results in insertion of a different amino acid into a protein and the appearance of an altered phenotype. Mutations result from three types of molecular changes:
(1) The first type is the base substitution. This occurs when one base is inserted in place of another. It takes place at the time of DNA replication, either because the DNA polymerase makes an error or because a mutagen alters the hydrogen bonding of the base being used as a template in such a manner that the wrong base is inserted. When the base substitution results in a codon that simply causes a different amino acid to be inserted, the mutation is called a missense mutation; when the base substitution generates a termination codon that stops protein synthesis prematurely, the mutation is called a nonsense mutation. Nonsense mutations almost always destroy protein function.
(2) The second type of mutation is the frameshift mutation. This occurs when one or more base pairs are added or deleted, which shifts the reading frame on the ribosome and results in incorporation of the wrong amino acids “downstream” from the mutation and in the production of an inactive protein.
(3) The third type of mutation occurs when transposons or insertion sequences are integrated into the DNA. These newly inserted pieces of DNA can cause profound changes in the genes into which they insert and in adjacent genes.
Mutations can be caused by chemicals, radiation, or viruses. Chemicals act in several different ways.
(1) Some, such as nitrous acid and alkylating agents, alter the existing base so that it forms a hydrogen bond preferentially with the wrong base (e.g., adenine would no longer pair with thymine but with cytosine).
(2) Some chemicals, such as 5-bromouracil, are base analogues, since they resemble normal bases. Because the bromine atom has an atomic radius similar to that of a methyl group, 5-bromouracil can be inserted in place of thymine (5-methyluracil). However, 5-bromouracil has less hydrogen-bonding fidelity than does thymine, and so it binds to guanine with greater frequency. This results in a transition from an A-T base pair to a G-C base pair, thereby producing a mutation. The antiviral drug iododeoxyuridine acts as a base analogue of thymidine.
(3) Some chemicals, such as benzpyrene, which is found in tobacco smoke, bind to the existing DNA bases and cause frameshift mutations. These chemicals, which are frequently carcinogens as well as mutagens, intercalate between the adjacent bases, thereby distorting and offsetting the DNA sequence.
X-rays and ultraviolet light can cause mutations also.
(1) X-rays have high energy and can damage DNA in three ways: (a) by breaking the covalent bonds that hold the ribose phosphate chain together, (b) by producing free radicals that can attack the bases, and (c) by altering the electrons in the bases and thus changing their hydrogen bonding.
(2) Ultraviolet radiation, which has lower energy than X-rays, causes the cross-linking of the adjacent pyrimidine bases to form dimers. This cross-linking (e.g., of adjacent thymines to form a thymine dimer) results in inability of the DNA to replicate properly.
Certain viruses, such as the bacterial virus Mu (mutator bacteriophage), cause a high frequency of mutations when their DNA is inserted into the bacterial chromosome. Since the viral DNA can insert into many different sites, mutations in various genes can occur. These mutations are either frameshift mutations or deletions.
Conditional lethal mutations are of medical interest because they may be useful in vaccines (e.g., influenza vaccine). The word conditional indicates that the mutation is expressed only under certain conditions. The most important conditional lethal mutations are the temperature-sensitive ones. Temperature-sensitive organisms can replicate at a relatively low, permissive temperature (e.g., 32°C) but cannot grow at a higher, restrictive temperature (e.g., 37°C). This behavior is due to a mutation that causes an amino acid change in an essential protein, allowing it to function normally at 32°C but not at 37°C because of an altered conformation at the higher temperature. An example of a conditional lethal mutant of medical importance is a strain of influenza virus currently used in an experimental vaccine. This vaccine contains a virus that cannot grow at 37°C and hence cannot infect the lungs and cause pneumonia, but it can grow at 32°C in the nose, where it can replicate and induce immunity.
TRANSFER OF DNA WITHIN BACTERIAL CELLS
Transposons transfer DNA from one site on the bacterial chromosome to another site or to a plasmid. They do so by synthesizing a copy of their DNA and inserting the copy at another site in the bacterial chromosome or the plasmid. The structure and function of transposons are described in Chapter 2, and their role in antimicrobial drug resistance is described in Chapter 11. The transfer of a transposon to a plasmid and the subsequent transfer of the plasmid to another bacterium by conjugation (see later) contributes significantly to the spread of antibiotic resistance.
Transfer of DNA within bacteria also occurs by programmed rearrangements (Figure 4–1). These gene rearrangements account for many of the antigenic changes seen in Neisseria gonorrhoeae and Borrelia recurrentis, the cause of relapsing fever. (They also occur in trypanosomes, which are discussed in Chapter 52.) A programmed rearrangement consists of the movement of a gene from a silent storage site where the gene is not expressed to an active site where transcription and translation occur. There are many silent genes that encode variants of the antigens, and the insertion of a new gene into the active site in a sequential, repeated programmed manner is the source of the consistent antigenic variation. These movements are not induced by an immune response but have the effect of allowing the organism to evade it.
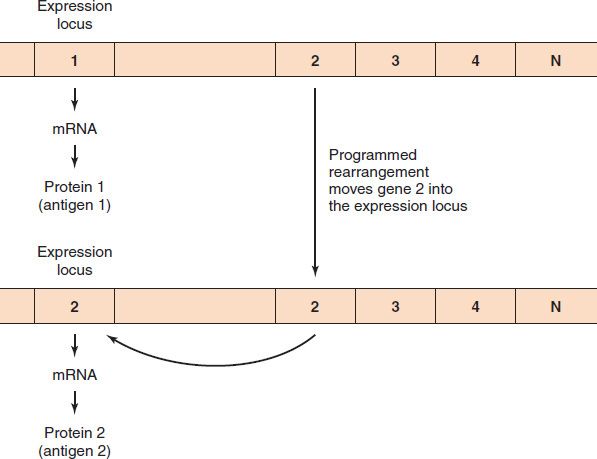
FIGURE 4–1 Programmed rearrangements. In the top part of the figure, the gene for protein 1 is in the expression locus and the mRNA for protein 1 is synthesized. At a later time, a copy of gene 2 is made and inserted into the expression locus. By moving only the copy of the gene, the cell always keeps the original DNA for use in the future. When the DNA of gene 2 is inserted, the DNA of gene 1 is excised and degraded.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


