Chapter 50 Antifungal Agents
| Abbreviations | |
|---|---|
| ABLC | Amphotericin B lipid complex |
| AIDS | Acquired immunodeficiency syndrome |
| CSF | Cerebrospinal fluid |
| GI | Gastrointestinal |
| IV | Intravenous |
Therapeutic Overview
| Therapeutic Overview |
|---|
| Cutaneous and Subcutaneous Mycoses |
| Treat with dermatological preparations, occasionally systemic agents |
| Epidermophyton species |
| Microspora species |
| Sporothrix species |
| Trichophyton species |
| Systemic Mycoses |
| Difficult to treat; available drugs often cause deleterious side effects; often need long-term therapy |
| Aspergillus species |
| Candida species |
| Blastomyces dermatitidis |
| Cryptococcus neoformans |
| Coccidioides immitis |
| Fusarium species |
| Histoplasma capsulatum |
| Paracoccidioides brasiliensis |
| Mucormycosis |
broad-spectrum antibacterial agent use). Many fungal infections are superficial and primarily annoying. Others are systemic and can be life-threatening, particularly in patients with compromised defenses, such as those receiving immunosuppressive drugs. The toxicity of many antifungal drugs limits their use, and unfortunately there are few agents useful in treating systemic fungal infections.
Mechanisms of Action
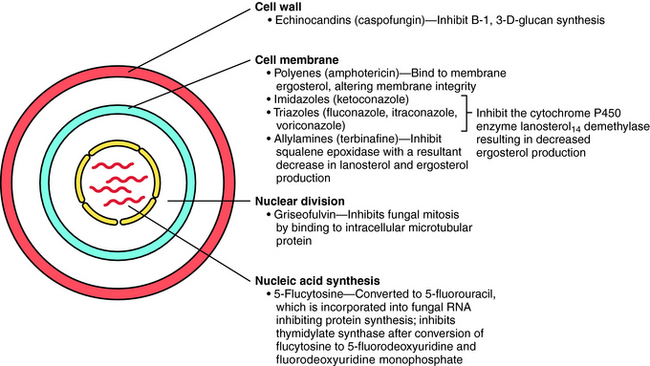
The principal antifungal drugs are the polyenes, azoles, allylamines, echinocandins, and others including flucytosine and griseofulvin. The sites of action of these agents are shown in Figure 50-1.
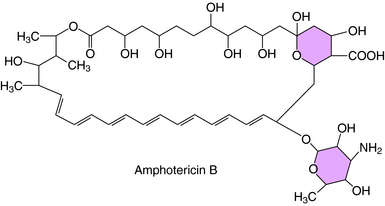
The polyene (i.e., multiple double bonds) drugs are macrocyclic lactones that contain a hydrophilic hydroxylated portion and a hydrophobic conjugated double bond portion. The structure of amphotericin B, the most widely used polyene antifungal drug, is shown in Figure 50-2.
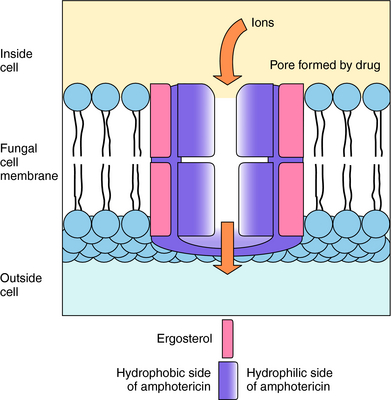
Polyenes act by binding to sterols in the cell membrane and forming channels, allowing K+ and Mg++ to leak out. The polyenes become integrated into the membrane to form a ring with a pore in the center approximately 0.8 nm in diameter. K+ leaks out through these pores, followed by Mg++, and with the loss of K+, cellular metabolism becomes deranged (Fig. 50-3). It is thought that derangement of the membrane alters activity of membrane enzymes. The principal sterol in fungal membranes, ergosterol, has a higher affinity for polyenes than does cholesterol, the principal sterol of mammalian cell membranes. Therefore the polyenes show greater activity against fungal cells than mammalian cells, and fungi that lack ergosterol are not susceptible to amphotericin B.
The structures of some of the principal azole antifungal agents are shown in Figure 50-4. Ketoconazole, miconazole, clotrimazole, and econazole are available, as are the newer agents fluconazole, itraconazole, and voriconazole.
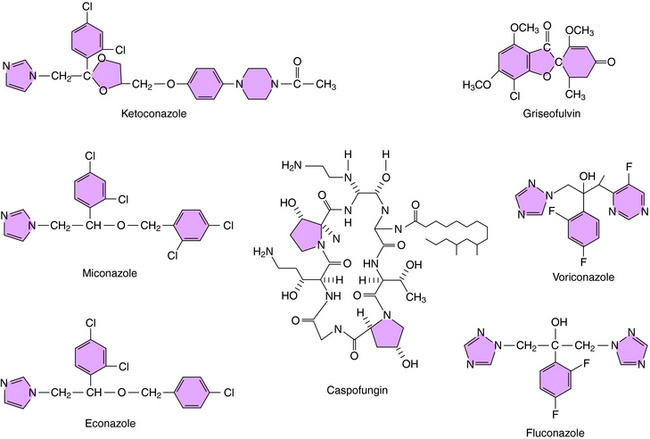
Caspofungin is the first echinocandin compound to gain approval for use in the United States. It blocks production of B-1,3-D-glucans, the major structural component of the fungal cell wall, by inhibition of glucan synthesis. Fungi have been shown to develop in vitro resistance to echinocandins through mutations in genes coding for the target enzymes.
Flucytosine is transported into susceptible fungi by a permease system for purines. The drug is then deaminated by cytosine deaminase to 5-fluorouracil. Because cytosine deaminase is not present in mammalian cells, the drug is not activated in humans. Fluorouracil in turn is converted by uridine phosphate pyrophosphorylase and other enzymes to 5-fluoro-2’-deoxyuridine 5’-monophosphate, which inhibits thymidylate synthase and interferes with deoxyribonucleic acid synthesis (Chapter 54).
Fungi can be resistant to flucytosine because they lack a permease, have a defective cytosine deaminase, or have a low concentration of the uridine monophosphate pyrophosphorylase. Whether the faulty ribonucleic acid produced by incorporation of fluorouracil contributes to its action is unclear.
Whether griseofulvin is fungicidal or fungistatic is not established. It enters susceptible fungi by an energy-dependent transport system and inhibits mitosis. It binds to the microtubules that form the mitotic spindle and blocks the polymerization of tubulin into microtubules. It also binds to a microtubule-associated protein, although the role of this protein is not known. The binding site for griseofulvin on tubulin differs from that of colchicine and the plant alkaloids. This effect on microtubule assembly probably explains the morphological changes, such as curling, that are observed in the fungi. Mechanisms of resistance are unknown, but may stem from decreased uptake of the drug. The structure of griseofulvin is shown in Figure 50-4.
Pharmacokinetics
Pharmacokinetic parameters for the antifungal drugs are summarized in Table 50-1.
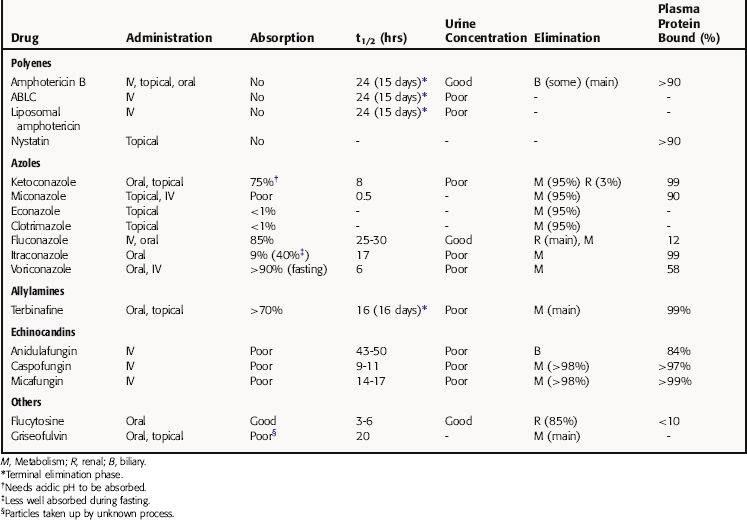
The principal pathway for amphotericin B elimination is not known. Some is excreted by the biliary route, and only 3% is eliminated in urine. Renal dysfunction does not affect plasma concentrations, and amphotericin B is not removed by hemodialysis.



