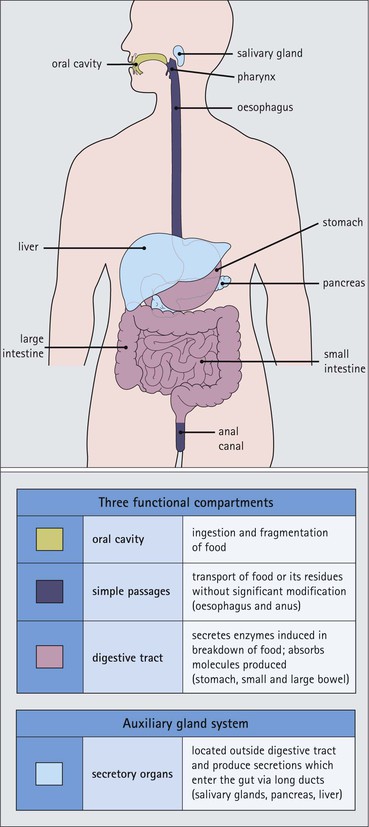
The alimentary tract is best considered as a muscular tube lined internally by an epithelium that varies in structure according to specialized functions required at particular sites along its length; with a few local variations, the structure of the musculature is similar throughout. The function of the alimentary tract is to take in raw food material and to fragment it into small portions. These are then acted upon by a series of secretions, mainly enzymes, which convert the large molecules into smaller molecules, thus permitting their absorption into the blood and lymph circulation. The small molecules are mainly amino acids, small peptides, carbohydrates, sugars and lipids, which are transported by the blood and lymph mainly to the liver, where they are used as the building blocks in the synthesis of essential proteins, carbohydrates and lipids. The alimentary tract can be divided into three functional components, with an additional auxiliary gland system The three functional components are the oral cavity, the simple transport passages and the digestive tract (Fig. 11.1). The oral cavity is the area where food is ingested, broken up into smaller fragments by the action of the teeth, and softened by secretions in the form of saliva from the salivary gland. The food material is moved around in the oral cavity by movement of the jaws and tongue to facilitate fragmentation. The moistened and fragmented bolus of food is then transferred by deglutition (swallowing) to the first of the simple passages, the oesophagus. The simple transport passages have no function other than to act as a contractile conduit to pass semisolid material from one area to another. The first simple transport passage is the pharynx, through which food from the mouth passes into the oesophagus which then transfers the moistened food bolus to the first part of the digestive tract, the stomach, before major digestion begins. The oesophagus is long, and the food bolus is forced along it by smooth muscle action in peristalsis, under involuntary nervous control. The other simple transport passage is the anal canal at the end of the digestive tract. This transports semisolid undigested waste material (faeces) from the end of the digestive tract to the exterior. The initial stimulus to transport is involuntary as a result of increasing distension of the rectum, but the timing of the evacuation of faeces can (usually) be controlled by voluntary (skeletal) muscle in an external sphincter. For semisolid material to pass easily through these simple transport passages, it is important that it is lubricated by mucus. For movement through the oesophagus, the mucus is incorporated into the food bolus in the mouth, having been secreted by mucous glands. In the anus, the faeces are lubricated by mucus secreted by the goblet cells in the colonic epithelium. The digestive tract proper comprises the stomach, the small intestine and large intestine (colon and rectum). The stomach acts as a reservoir where the ingested fragmented food is held up by a sphincter until the acid and enzymatic secretions of the stomach mucosa have broken up the fragmented food into a semiliquid slurry (chyme), which then passes through the sphincter into the small intestine. Here, the process of digestion is continued by a combination of enzymes and other chemicals secreted by the small intestine itself and by the auxiliary glands, the liver and pancreas, the secretions of which enter the small intestine via ducts. In addition to this digestion function, the main function of the small intestine is absorption of the breakdown products of food digestion. The liquid food residue passes from the small intestine into the large intestine, where the majority of the fluid content is reabsorbed until the waste material is converted into semisolid waste material, lubricated for easy passage through the anal canal by mucus secreted by goblet cells in the colon and rectum. All the components of the alimentary tract are structurally specialized to carry out their specific functions. The auxiliary gland systems contributing to the function of the alimentary tract are the salivary glands, the liver and the pancreas, all of which pass their secretions into the main alimentary tract through ducts. The salivary glands are discussed on page 197, the pancreas on page 214, and the liver, which has many important functions other than those associated with digestion, merits a chapter of its own (Chapter 12). The oral cavity is lined throughout by stratified squamous epithelium but contains many highly specialized structures The mouth is lined by stratified squamous epithelium, the underlying submucosa containing varying numbers of salivary glands, which can secrete both a serous and a mucous fluid. Skeletal muscle fibres are common in the deeper layers and are responsible for altering the size and shape of the cavity and for moving food; skeletal muscle fibres form the bulk of the tongue, and are also numerous and important in the cheeks. In some areas, the deep tissues of the oral cavity consist of bone, either simple plates of bone (as in the hard palate) or modified bone (teeth). The immovable hard palate produces a rigid structure against which the tongue can move, and the teeth, which are embedded in bony supports (the mandibles and maxillae), are the main tools for fragmenting ingested food. The lips are covered by squamous epithelium and contain glands and underlying muscle The oral orifice is bounded by the lips, the external aspects of which are covered by hair-bearing skin with sebaceous glands and eccrine sweat ducts. Between the hair-bearing outer surface and the moist, fluid-bathed inner surface, there is a transitional zone known as the vermilion because of its pinkish-red appearance. Here, the epithelium is non-keratinizing stratified squamous with a prominent rete ridge system (see Chapter 18) and the papillae between the epithelial downgrowths contain prominent blood vessels, which are responsible for the pinkish-red colour. The inner surface of the lips is lined by a non-keratinizing stratified squamous epithelium with a less well-developed rete ridge system, and small clumps of salivary tissue disgorge their secretions on to its surface through short ducts. In addition, occasional sebaceous glands (Fordyce’s spots), which are particularly common near the angles of the mouth, open directly on to the mucosal surface, rather than into a hair follicle as in the skin. In the deeper parts of the lips, bundles of striated muscle fibres (orbicularis oris muscle) are arranged mainly in a concentric manner around the oral orifice. This muscle is responsible, among other things, for opening and closing the oral orifice. The cheeks are lined internally by squamous epithelium and contain glands and deep muscle The cheeks are lined by thick non-keratinizing squamous epithelium, the cells of which are often rich in glycogen. Areas of keratinization are common, usually as a result of chronic friction from ill-fitting dentures or from persistent cheek biting. The submucosa contains minor salivary glands (buccal glands) and occasional sebaceous glands (Fordyce’s spots), whereas the deep tissues contain the skeletal muscle fibres of the cheek muscles (buccinator). The palate is rich in salivary gland tissue (palatine salivary glands) The surface epithelium of the palate is non-keratinizing, stratified squamous epithelium with, in the hard palate, a prominent rete ridge pattern, which reflects the frictional shearing stresses this area is subjected to by food during mastication. Beneath the salivary gland tissue the submucosa is firmly tethered to the periosteum of the palatal bone plate. The oral surface of the soft palate is similarly covered with non-keratinizing stratified squamous epithelium, which extends on to its free posterior edge, where there is a transition to the ciliated columnar epithelium covering the nasal surface. The floor of the mouth contains salivary glands The floor of the mouth is covered by thin non-keratinized stratified squamous epithelium, which is continuous with that of the ventral surface of the tongue. The floor of the mouth is also rich in salivary gland tissue, with many minor salivary glands in the floor of the mouth (minor sublingual glands) and larger glands situated on either side of the midline frenulum of the ventral surface of the tongue (major sublingual glands, see p. 197). The tongue is muscular and covered in squamous epithelium The tongue is a highly muscular organ that protrudes upwards and forwards into the oral cavity from its floor. The ventral surface of the tongue is covered by thin non-keratinizing stratified squamous epithelium, continuous with that of the floor of the mouth. In contrast, the dorsal surface, which is commonly in contact with the hard palate during feeding, talking and at rest, is covered with a thick keratinizing stratified squamous epithelium, which shows considerable specialization. The upper surface of the tongue is divided into two main zones The dorsal surface of the tongue is divided into an anterior two-thirds and a posterior one-third. The two parts are separated by a V-shaped line of 6–10 dome-shaped protrusions, the ‘circumvallate papillae’ (Fig. 11.2). Circumvallate papillae appear as flattened domes, the bases of which are depressed below the dorsal surface. Each circumvallate papilla is surrounded by a narrow, moat-like channel, in the epithelium of which are numerous taste buds. These taste buds are thought to detect bitter taste. Small salivary glands discharge their secretions into the channels (Fig. 11.3). The posterior third of the tongue is characterized by the presence of lymphoid tissue Low smooth dome-shaped elevations of the covering epithelium of the posterior third of the tongue are due to lymphoid tissue (the lingual tonsillar tissue) in the submucosa (Fig. 11.4). This lymphoid tissue is part of the mucosa-associated lymphoid tissue system (MALT, see p. 140) protecting (with the palatine tonsils and the pharyngeal adenoids) the oral portal of entry. Lymphocytes are numerous within the overlying non-keratinizing stratified squamous epithelium, which extends down into the lymphoid tissue as narrow clefts. Small salivary glands open into the bottom of the narrow clefts, becoming more prominent and numerous near the line of circumvallate papillae. The surface epithelium of the anterior two-thirds of the tongue is raised in a series of elevations called papillae The three types of papillae in man are circumvallate (described above), filiform and fungiform. Filiform papillae are the most numerous papillae and are found all over the dorsum of the anterior two-thirds of the tongue. They are tall, narrow and pointed (Fig. 11.5) and are keratinized, particularly at their tips. Filiform papillae contain no identifiable taste buds. Fungiform papillae (see Fig. 11.5) are scattered apparently randomly among the filiform papillae on the dorsal surface of the tongue, and have a mushroom shape. Taste buds are present in their covering epithelium, those at the anterior tip of the tongue detecting sweet taste, and those just behind the tip and some way along the lateral borders detecting salty taste. Taste buds are specialized sensory organs located in the epithelium of the tongue Each taste bud occupies the full thickness of the epithelium and comprises pale-staining spindle-shaped cells in an oval cluster (Fig. 11.6). The luminal surfaces of the cells open into a small defect in the epithelium, the taste pore, and each cell bears a number of microvilli. Ultrastructurally, some of the spindle-shaped cells have synaptic vesicles and are associated with small afferent nerve fibres; these are the taste receptor cells. Other cells with more electron-dense cytoplasm and scanty secretory granules near the luminal surface are thought to act mainly as supporting sustentacular cells, but may also secrete glycosaminoglycans into the taste pore. There are also cells resembling the taste receptor cells but lacking the synaptic vesicles and afferent nerve connections. The turnover of these cells is rapid (every 10–14 days), so there is a population of small rounded stem cells at the base of each taste bud from which the other cell types derive. The taste buds of the dorsal tongue detect only acid, sweet, bitter and salty, and provide early warning that food may be unpalatable. Appreciation of more subtle flavours depends on the smell receptors in the nose (see p. 168). Skeletal muscle in the tongue is arranged in many directions The musculature of the tongue comprises a complex pattern of skeletal muscle fibres. These fibres run in bands longitudinally, vertically, transversely and obliquely, with a variable amount of adipose tissue in between (Fig. 11.7). This arrangement gives the tongue great mobility to manipulate food around the mouth for efficient fragmentation, and for moving fragmented food backwards prior to swallowing; it also provides the fine control of tongue movement that is essential for speech. Abundant islands of salivary tissue are present in the submucosa between the muscular core of the tongue and the surface epithelium in the region of the junction between the posterior one-third and the anterior two-thirds. Some of the deeper salivary gland collections extend down into the superficial parts of the muscular zone. The main task of the oral cavity, the fragmentation of large food masses, is performed by the teeth, which are hard, heavily mineralized structures embedded in the raised alveolar ridges of the maxilla and mandible. Teeth are arranged so that the free surface of those embedded in the mandible (lower teeth) oppose and contact those in the maxilla (upper teeth), allowing food material to be trapped between them. The anterior teeth (incisor and canine teeth) have narrow, chisel-shaped or pointed free edges for chopping food into medium-sized pieces, whereas the posterior teeth (premolars and molars) have broader, flatter, free surfaces for grinding medium-sized food pieces into smaller fragments. The mandible is joined to the body of the skull by the temporomandibular joint, which permits the mandible to slide backwards and forwards, and from side to side, thus aiding the grinding of food between the broad surfaces of the opposing molar teeth. Each tooth can be divided into two anatomical components: the crown and the root The tooth is divided into two structural regions: • The crown protrudes into the oral cavity • The root is embedded in the bone of the mandible or maxilla. The junction between the crown and the root is called ‘the neck’. The mature tooth has five components: the central pulp cavity, dentine, enamel, cementum and periodontal ligament (Fig. 11.8). The centre of each tooth comprises the pulp, which contains nutrient vessels and nerves The pulp cavity is the soft central core of the tooth and contains collagen and fibroblasts embedded in an acellular matrix. The cavity approximates in shape that of the tooth as a whole and its acellular matrix is composed of glycosaminoglycans. Through the pulp cavity run the blood vessels that nourish the odontoblasts (see below) and the nerve twigs that provide dental sensation. These vessels and nerves enter and leave through a small apical foramen at the tip of the root. The pulp cavity is narrow throughout most of the root (the root canal), but is expanded in the neck and crown (the pulp chamber). Its outer surface is lined by odontoblasts, which continually produce dentine. As dentine is progressively laid down, the pulp cavity diminishes in size. The specialized mineralized matrix of the tooth is called dentine and is secreted by odontoblasts Dentine is composed of mineral salts and organic material. The mineral salts are calcium salts in the form of crystalline hydroxyapatite (about 70–80%) arranged as long, hollow parallel tubules, the dentinal tubules. Within these, run the organic material in the form of fine cytoplasmic processes of the odontoblasts, and the type I collagen fibres and glycosaminoglycans that they produce (about 20–30%). Dentine is initially laid down by odontoblasts as a glycosaminoglycan matrix in which collagen fibres are linearly arranged. This non-mineralized predentine is synthesized by odontoblasts located at the outer limits of the pulp cavity (Fig. 11.9). In man, the odontoblast cytoplasmic processes extend only 25–50% of the full length of the dentinal tubule, thus dentine close to the dentine–enamel border appears to contain empty tubules. In life, these empty tubules may contain fluid, which is lost during tissue processing. As odontoblasts progressively synthesize new predentine at the inner surface of the pulp cavity (Figs 11.9, 11.10), the cavity slowly decreases in size throughout life. Dentinogenesis commences with the formation of predentine by odontoblasts In predentine, randomly scattered collagen fibres (type I collagen) produced by the odontoblasts are embedded in an extracellular matrix of phosphoprotein and glycosaminoglycans (mainly chondroitin-6-sulfate). Mineralization is initiated by the discharge of the matrix vacuoles from the odontoblast process and its branches running through the predentine layer. Some time after its formation, predentine becomes mineralized at its border with previously mineralized dentine; close to this predentine–dentine border the collagen fibres of predentine become more numerous and tightly packed. The dentine lining the dentinal tubules (peritubular dentine) is particularly compact and heavily mineralized (Fig. 11.11). Enamel is the hardest material in the body Enamel is composed almost entirely of the mineral hydroxyapatite (Ca10(PO4)6(OH)2), which is arranged in tightly packed hexagonal enamel rods or prisms (Fig. 11.12) about 4 µm in diameter, although some may measure up to 8 µm. Each enamel rod extends through the full thickness of the enamel. The small interstices between adjacent rods are occupied by hydroxyapatite crystals. A small amount of organic matrix (protein and polysaccharide) represents the remnants of the matrix synthesized and excreted by the enamel-producing cells, ‘the ameloblasts’, prior to mineralization of the enamel. Enamel is formed during tooth development by the ameloblasts Ameloblasts degenerate when the tooth erupts, after which time the enamel cannot be replaced by new synthesis. In the developing tooth (see Fig. 11.17), the functioning ameloblast is a tall, narrow cell, with its base attached to the cells of the stratum intermedium (Fig. 11.13). The nucleus is located basally and basal cytoplasm contains abundant mitochondria. The supranuclear cytoplasm contains a large, active Golgi complex and abundant rough endoplasmic reticulum, together with microtubules, which are predominantly longitudinally arranged, and secretory vacuoles which become larger and more numerous near the upper pole. At the upper pole, the cell elongates into a single large Tomes’ process, and forms a fringe of smaller processes around its neck. The Tomes’ process contains numerous microtubules and large numbers of secretory vacuoles. The rough endoplasmic reticulum synthesizes various proteins and glycoproteins (including amelogenin and enamelin), which form the organic matrix of enamel (pre-enamel) and are packaged by the Golgi complex into secretory vacuoles. These then move into the Tomes’ process and the small neck processes, discharging their contents on to the surface. Mineralization of the matrix proteins by hydroxyapatite occurs almost instantaneously, producing small enamel crystallites and, with progressive mineralization, the enamel rods or prisms. The compact structured enamel prisms are probably derived from the surface of the main Tomes’ process, whereas the small amount of less compact interprism enamel, which has a larger organic matrix component, is probably derived from the small neck processes. Enamel covers the dentine only in the region of the exposed crown (Fig. 11.14); in the root, the dentine is covered by cementum (see Figs 11.8, 11.15). Cementum is a bone-like tissue; it is calcified and contains collagen A thin layer of cementum covers the root of the tooth, being thin, compact and acellular (acellular cementum) over the upper region, but thicker and containing lacunae and cementocytes (cellular cementum) lower down (see Fig. 11.15). Cementocytes resemble osteocytes (see Chapter 13) and remain viable throughout life, being nourished through canaliculi which link the lacunae. They can become activated to produce new cementum when required. In addition to the cementocytes, which are scattered throughout the cellular cementum, there is a layer of cells called ‘cementoblasts’, which are similar to the actively synthetic osteoblasts of bone (see Fig. 13.17). Cementoblasts lie against the surface of the periodontal ligament and probably produce most new cementum by appositional deposition. The periodontal ligament is a suspensory ligament tethering the tooth in the bony alveolar socket of the mandible or maxilla The periodontal ligament is composed of dense collagen and fibrocytes, with the fibres running across the gap between the cementum of the tooth and the bone of the alveolar socket (Fig. 11.16). As the collagen fibres are embedded in a ground substance, this ligament also acts as a shock-absorber, as well as permitting limited movement of the tooth within the bony socket. At its cemental and alveolar limits, some of the collagen fibres are inserted into the cementum and bone. The alveolar bone at the inner margin of the socket is composed of woven bone rather than compact lamellar bone. Above the upper limit of the alveolar bone, the fibres of the periodontal ligament become the gingival periodontal fibres and blend with the submucosa of the gingivae. Dental Caries and Periodontal Disease The most common disorders of teeth are: Dental caries results when bacterial plaque produces acid, which dissolves the calcium hydroxyapatite of the enamel. Such focal decalcification may progress until the erosion involves the deeper dentine, from where the acids and bacteria can advance more rapidly down the dentinal tubules to the pulp cavity, causing tooth pain. Continued bacterial damage can produce a tooth abscess in the soft pulp. Destruction of the pulp cavity and its contained blood vessels leads to death of the odontoblast layer and eventual death of the tooth. Periodontal disease is caused by the accumulation of bacterial plaque (calcified food and bacterial debris) in the gingival sulcus. This excites inflammation in the adjacent gum which gradually forces the gingiva away from the tooth, widening and damaging the gingival sulcus; deep periodontal pockets form in which food particles and bacteria become trapped. Bacterial proliferation then leads to further inflammation of the gum (gingivitis) and the periodontal ligament (periodontitis). Persistent periodontitis destroys the periodontal ligament and the tooth becomes loose in its socket. Teeth develop from both ectoderm and mesoderm Teeth develop from both ectoderm and from mesenchyme of mesodermal origin, the ectodermal component being the enamel derived from the enamel organ and the mesenchyme producing the rest of the tooth. The enamel organ originates as a cellular downgrowth of the oral epithelium, initially in the form of a cap-shaped epithelial tooth bud connected to the overlying oral epithelium by the dental lamina. Beneath the epithelial tooth bud, a condensation of mesenchyme contributes to the rest of the tooth. The stages in tooth formation are summarized in Figure 11.17. The gingiva (gum) is the portion of oral mucosa covering the alveolar bone ridge surrounding the tooth The gingival mucosa is continuous with the alveolar mucosa covering the rest of the bone. A small sulcus lies between the tooth and the bulk of the gingiva. The sulcal epithelium is thin stratified squamous, whereas that on the outer surface of the gingiva is thick and keratinized, with a prominent rete ridge pattern. The submucosa in the region of the gingival sulcus is commonly infiltrated by chronic inflammatory cells and also contains gingival periodontal fibres, which are an extension of the upper limit of the periodontal ligament. There are three main salivary glands and many minor glands The mouth receives secretions from salivary gland tissue located both inside and outside the mouth. These glands may contain mucus-secreting cells, serous cells or a mixture of both. The serous glands secrete a watery solution containing enzymes (e.g. amylase, lysozyme), IgA secretory piece and lactoferrin, an iron-binding compound. The largest salivary glands are located outside the mouth and transfer their secretions into the oral cavity by long ducts; smaller glands, sited mainly in the submucosa of the oral lining, are less well-defined and empty their secretions into the mouth by short ducts. The main salivary glands are the submandibular, the parotid and the large sublingual glands. The submandibular salivary gland secretes a mixed serous/mucous product The submandibular glands are roughly ovoid in shape and are situated on either side of the neck just below the mandible. Their ducts open into the floor of the mouth, one on each side of the frenulum of the tongue. The submandibular glands are typical mixed glands containing both serous and mucous elements, with serous elements predominating. The secretory acini are composed mainly of epithelial cells, which are responsible for the serous secretion, and are plump cells filled with prominent, purplish-staining zymogen granules. The mucus-secreting cells are pale-staining with abundant clear cytoplasm, and often form blind-ending tubules, the blind end bearing a demilune or crescent of zymogen-rich serous cells (Fig. 11.18). The secretory acini empty into intercalated ducts, which are lined by cuboidal or low columnar epithelium and merge to form intralobular ducts. These are characterized by tall columnar epithelium, which stains pale pink and shows a characteristic striated pattern (see Fig. 3.23) of basal cytoplasm. Hence, intralobular ducts are commonly called ‘striated ducts’. Intralobular duct epithelium is biochemically active and modifies the concentration and content of the fluids produced by the secretory acini. The intralobular striated ducts fuse to form larger interlobular ducts which are lined by a non-striated, often pseudostratified, epithelium. Interlobular ducts then join to form the major ducts, some of which (particularly the main submandibular duct) may have a ciliated epithelium. There is a network of myoepithelial cells between the epithelium and the basement membrane of the acini and much of the ductular system, contraction of the myoepithelial cells squeezing the secretion toward the major ducts. The parotid salivary glands secrete a serous product The parotid glands are situated below and in front of the pinna on each side of the face. They are flat and well encapsulated and the facial nerve runs through them, dividing them into superficial and deep portions. Their long ducts open into the oral cavity opposite the second upper molar tooth on each side. The parotids are composed entirely of serous glands rich in zymogen granules, with a variable amount of adipose tissue in the interstitium between parotid lobules (Fig. 11.19). The sublingual glands secrete a mainly mucous product The large sublingual glands are located in the floor of the mouth, one on either side of the frenulum of the tongue. Their short ducts open into the mouth near to, or with, the submandibular ducts. These glands are composed predominantly of mucous cells. There are numerous smaller groups of salivary gland tissue The oral cavity contains a large amount of salivary gland tissue scattered diffusely in the submucosa. The most important are: • Lingual glands in the submucosa and muscle layers of the dorsal surface of the tongue (see Fig. 11.7b) • Labial glands on the inner surface of the lips • Palatine glands in the submucosa of the soft and hard palates • Tonsillar glands in the mucosa associated with the palatine and pharyngeal tonsils The labial, sublingual, minor lingual and buccal glands are composed predominantly of mucous cells, but some serous cells may be present. The palatine and lateral lingual glands are entirely mucus secreting. The pharynx, oesophagus and anal canal are comparatively simple transport tubes through which ingested food is transmitted by peristalsis without undergoing significant metabolic change. They are muscular tubes lined internally by stratified squamous epithelium and some mucous glands which provide lubricating mucus. The pharynx is divided into oropharynx and nasopharynx The pharynx is located at the back of the mouth and transfers partly fragmented food from the oral cavity into the upper end of the oesophagus. It also connects the nasal system of air chambers and the upper end of the trachea. The opening of the mouth into the pharynx is the oropharynx, and the nasal opening is the nasopharynx. The eustachian tube from the middle ear (see Chapter 10, p. 167) opens into the pharynx on each side. The oropharynx and pharynx proper are lined by largely non-keratinizing stratified squamous epithelium. The nasopharynx is mainly lined by ciliated columnar epithelium but stratified squamous epithelium occurs at its lower end where it joins the oropharynx. The submucosa of the pharynx is well endowed with lymphoid tissue
Alimentary Tract
Introduction
Oral Cavity and its Contents
 Key Facts
Key Facts
 Key Facts
Key Facts
Teeth
Introduction
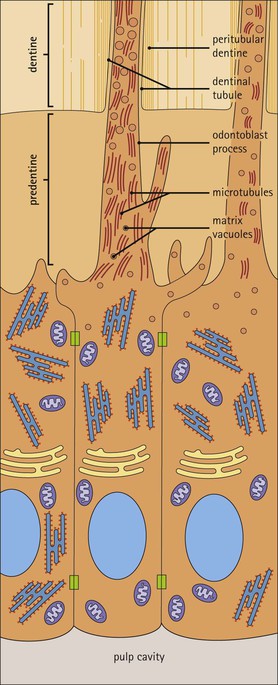
Dentinogenesis and Odontoblasts
Ameloblasts and Enamel Formation
Cementum and Periodontal Ligament
 Clinical Example
Clinical Example
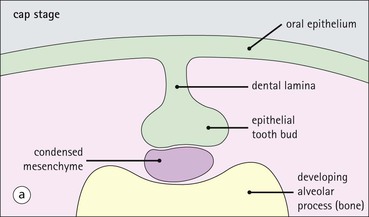
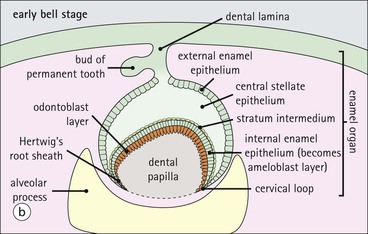
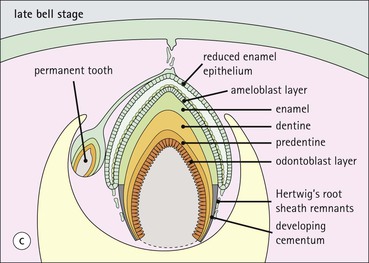
Tooth Development
The Gums
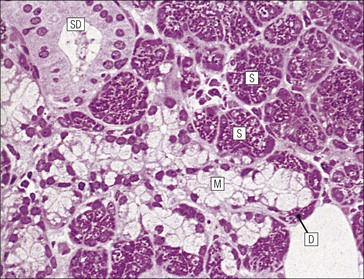
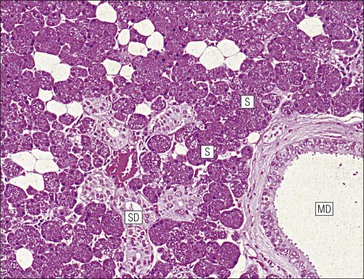
Salivary Glands
Transport Passages
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Alimentary Tract
Chapter 11
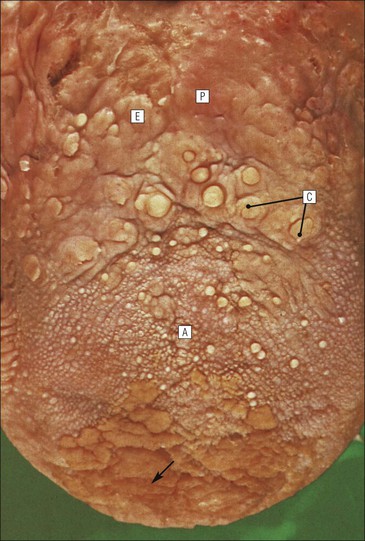
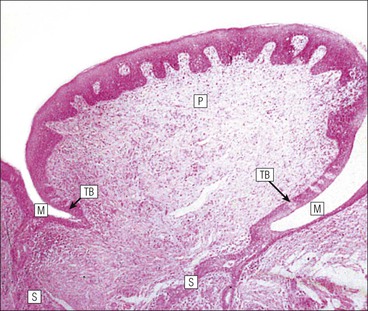
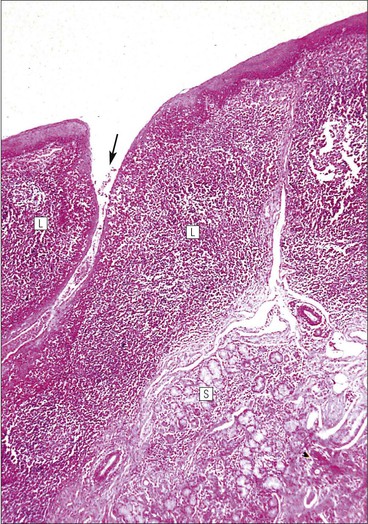
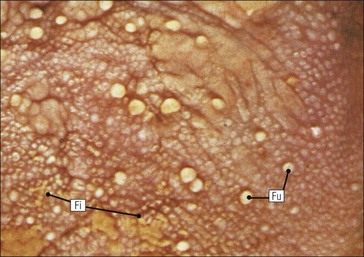
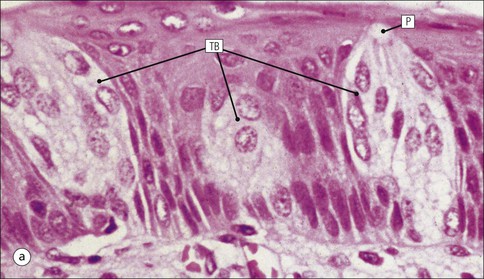
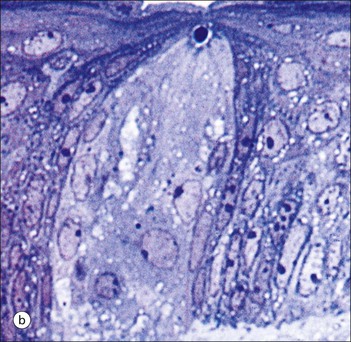
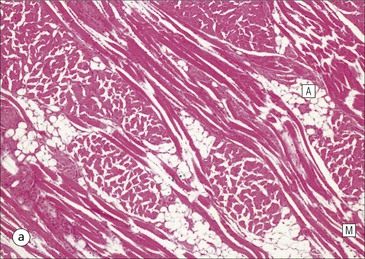
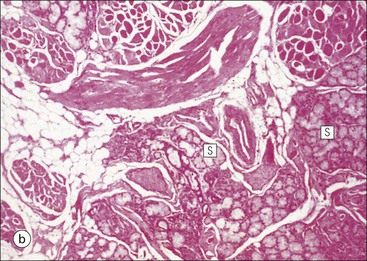
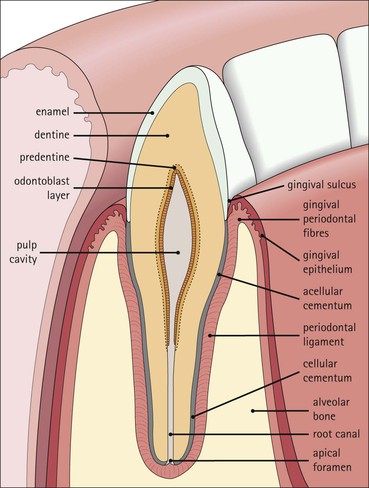
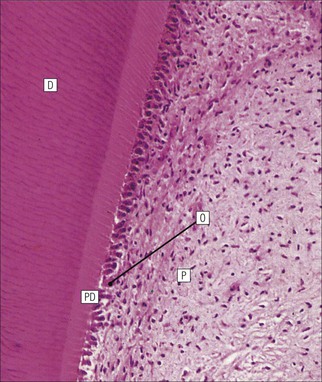
FIGURE 11.10 Odontoblasts and dentinogenesis. The functioning odontoblast is a tall, narrow cell, the base of which contacts cells and fibres of the pulp cavity (mainly fibroblasts and collagen). Its nucleus is basal and its cytoplasm is rich in mitochondria and rough endoplasmic reticulum. It has a large supranuclear Golgi complex. At the apex of the cell the cytoplasm is drawn out into a long odontoblast process which runs through the predentine and dentine layers in the dentinal tubule, while small side branches penetrate the predentine. The cytoplasm of the odontoblast process and its branches contains numerous microtubules as well as small matrix vacuoles, which contain Ca2+ and PO4– ions. The matrix vacuoles play a key role in the mineralization of the dentinal matrix. The mechanism is almost identical to that whereby matrix vesicles from osteoblasts produce mineralization of osteoid matrix to produce bone mineralization (see Fig. 13.20).
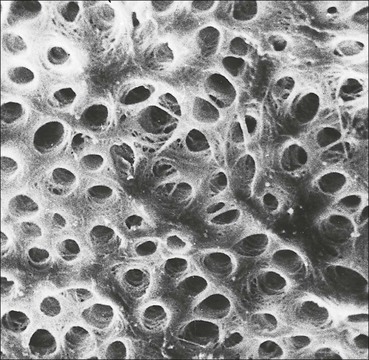
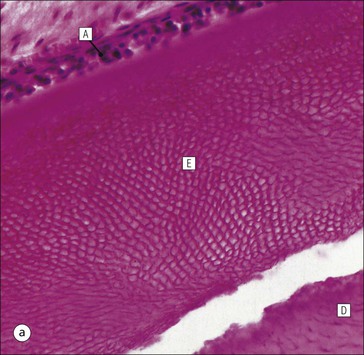

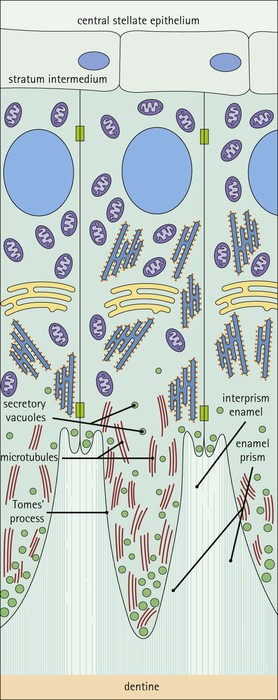

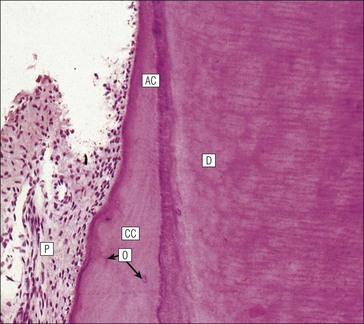
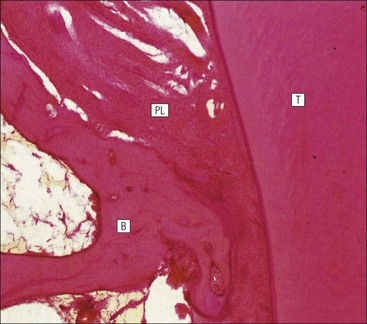
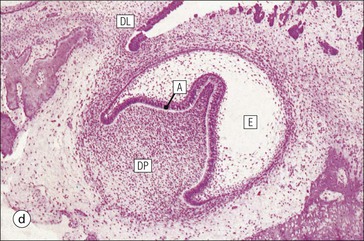
FIGURE 11.17 Tooth development. (a) Early stage of tooth formation from both ectodermal (epithelial) and mesenchymal components. (b) The cells of the epithelial tooth bud develop into the enamel organ, differentiating into a bell-shaped structure with a central core of loosely arranged stellate cells (stellate epithelium) and a peripheral layer of cuboidal or low columnar epithelium. The outer cell layer of the convex surface is the external enamel epithelium, and that of the concave surface the internal enamel epithelium. The internal enamel epithelium differentiates into an outer layer of tall columnar ameloblasts (see Fig. 11.13) and a two- to three-cell thick inner layer called the ‘stratum intermedium’. Where the external and internal enamel epithelium meet is called the ‘cervical loop’. An extension downward of cells of the external enamel epithelium forms the so-called ‘Hertwig’s root sheath’, which defines the final size of the tooth root, being later replaced by the cementum. In the concavity of the enamel organ the mesenchyme continues to condense to form the dental papilla, and a row of odontoblasts develops at its junction with the enamel organ, in contact with the ameloblast layer. In deciduous teeth, like the incisor shown here, the permanent tooth arises from a side-growth from the dental lamina. (c) Odontoblasts begin to produce predentine and this stimulates the production of enamel by the ameloblasts. Calcification of the predentine and pre-enamel begins almost immediately, and dentine and enamel continue to be laid down until the form of the tooth is complete. The dental papilla becomes enclosed by dentine to form the dental pulp. The non-ameloblast components of the enamel organ become much reduced and eventually atrophy. When enamel formation is complete, the ameloblasts degenerate to form a thin layer of irregular cells, which ultimately disappear when the tooth erupts. The cells of Hertwig’s root sheath begin to degenerate as the cementum is deposited by cementoblasts on the surface of the dentine of the tooth root. Partial development of the permanent tooth continues alongside the deciduous tooth in the same manner. (d) Micrograph of a resin section of a developing tooth at the early bell stage, showing the enamel organ (E), including the ameloblast layer (A), the dental papilla (DP) and the degenerating dental lamina (DL).