Urinary System
LANGUAGE OF SCIENCE
 Before reading the chapter, say each of these terms out loud. This will help you avoid stumbling over them as you read.
Before reading the chapter, say each of these terms out loud. This will help you avoid stumbling over them as you read.
atrial natriuretic hormone (ANH)
(AY-tree-al nay-tree-yoo-RET-ik HOR-mohn)
[William Bowman English anatomist]
[cortic- bark (cortex), -al relating to, nephro- kidney, -on unit]
[counter- against, -current flow]
distal convoluted tubule (DCT)
(DIS-tall KON-vo-LOO-ted TOO-byool)
[dist- distance, -al relating to, con- together, -volut- roll, tub- tube, -ul- little]
[filtr- strain, -ation process]
(gloh-MER-yoo-lar KAP-soo-lahr MEM-brayne)
glomerular filtration rate (GFR)
(gloh-MER-yoo-lar fil-TRAY-shun)
[glomer- ball, -ul- little, -ar relating to, filtr- strain, -ation process]
[glomer- ball, -ulus little] pl., glomeruli]
[Friedrich Gustave Henle German anatomist]
(juks-tah-gloh-MER-yoo-lar app-ah-RAT-us)
[juxta- near or adjoining, -glomer- ball, -ul- little, -ar relating to]
[juxta- near or adjoining, –glomer- ball, -ul- little, -ar relating to, cell storeroom]
(jux-tah-MED-oo-lair-ee NEF-ron)
[juxta- near or adjoining, -medulla- middle, -ary relating to, nephro- kidney, -on unit]
[macula spot, densa thick] pl., maculae densae
[mes- middle, -angi- vessel, -al relating to]
[myo- muscle, -gen- produce, -ic relating to]
(pair-ee-TOOB-yoo-lar KAP-i-lair-ee)
[peri- around, –tub- tube, -ul- little, -ar relating to, capill- hair, -ary relating to]
proximal convoluted tubule (PCT)
(PROK-si-mal KON-vo-LOO-ted TOO-byool)
[proxima- near, -al relating to, con- together, -volut- roll, tub- tube, -ul- little]
[re- back again, -absorp- swallow, -tion process]
[ren- kidney, -al relating to]
[ren- kidney, -al relating to]
[ren- kidney, -al relating to, corpus- body, -cle little]
[ren- kidney, -al relating to, cortex bark] pl., cortices
[ren- kidney, -al relating to, medulla middle] pl., medullae or medullas
[ren- kidney, -al relating to, pelvis basin]
[ren- kidney, -al relating to]
[sod- soda, -ium chemical ending, co- with, -trans- across, -port carry]
(TOOB-yoo-lar ree-ab-SORP-shun)
[tub- tube, -ul- little, -ar relating to, re- back again, -absorp- swallow, -tion process]
[tub- tube, -ul- little, -ar relating to, secret- separate, -tion process]
(toob-yoo-loh-glow-MER-yoo-lar)
[tub- tube, -ul- little, –glomer- ball, -ul- little, -ar relating to]
[ure- urine, -ter- agent or channel]
[ure- urine, -thr- agent or channel]
LANGUAGE OF MEDICINE
(ah-KYOOT gloh-mer-yoo-loh-neh-FRY-tis)
[acut- sharp, glomer- ball, -ul- little, -nephr- kidney, -itis inflammation]
[a- not, -ur- urine, -ia condition]
blood urea nitrogen (BUN) test
[ure- urine, nitro- soda, -gen produce]
(KRON-ik gloh-mer-yoo-loh-neh-FRY-tis)
[chron- time, -ic relating to, glomer- ball, -ul- little, -nephr-kidney, -itis inflammation]
[chron- time, -ic relating to, ren- kidney, -al relating to]
continuous ambulatory peritoneal dialysis (CAPD)
(AM-byoo-lah-tor-ee pair-i-toh-NEE-al dye-AL-i-sis)
[cyst- bag, -itis inflammation]
[dys- painful, -ur- urine, -ia condition]
(gloh-mer-yoo-loh-neh-FRY-tis)
[glomer- ball, -ul- little, -nephr-kidney, -itis inflammation]
[glyco- sweet (glucose), -ur- urine, -ia condition]
[hema- blood, -ur- urine, -ia condition]
[hemo- blood, -dia- through or between, –lysis loosening]
[hydro- water, -nephr- kidney, -osis condition]
[inter- between, -stit- stand, -al relating to, cyst- bag, -itis inflammation]
[litho- stone, -trips- pound, -y action]
[litho- stone, -trip- pound, -or agent]
[nephr- kidney, -itis inflammation]
[nephr- kidney, -ic relating to]
[neuro- nerves, -gen- produce, -ic relating to]
[olig- few or little, -ur- urine, -ia condition]
[poly- many, -ur- urine, -ia condition]
postinfectious glomerulonephritis
(post-in-FEK-shus gloh-mer-yoo-loh-neh-FRY-tis)
[pyel- renal pelvis, -nephr- kidney, -itis inflammation]
[py- pus, -ur- urine, -ia condition]
[ren- kidney, -al relating to, calculi little stone] pl., calculi
[ren- kidney, -al relating to, cell storeroom, carcino- cancer, -oma tumor]
[ren- kidney, -al relating to]
[ren- kidney, -al relating to, ptosis falling]
[stric- tighten, -ture condition]
[ur- urine, -emia blood condition]
[ur- urine, -(h)em- blood, -ic relating to, syn- together, -drome running or (race)course]
[ure- urine, -thr- agent or channel (urethra), -itis inflammation]
ANATOMY OF THE URINARY SYSTEM
Gross Structure
The principal organs of the urinary system are the kidneys, which process blood and form urine as a waste to be excreted—that is, removed from the body (Box 31-1). The excreted urine travels from the kidneys to the outside of the body via accessory organs: the ureters, urinary bladder, and urethra.
Box 31-1
 FYI
FYIExcretion
The urinary system’s chief function is to regulate the volume and composition of body fluids and excrete unwanted material, but it is not the only system in the body that is able to excrete unneeded substances.
The table below compares the excretory functions of several systems. Although all of these systems contribute to the body’s effort to remove wastes, only the urinary system can finely adjust the water and electrolyte balance to the degree required for normal homeostasis of body fluids.
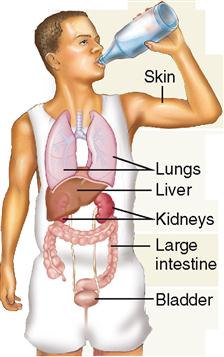
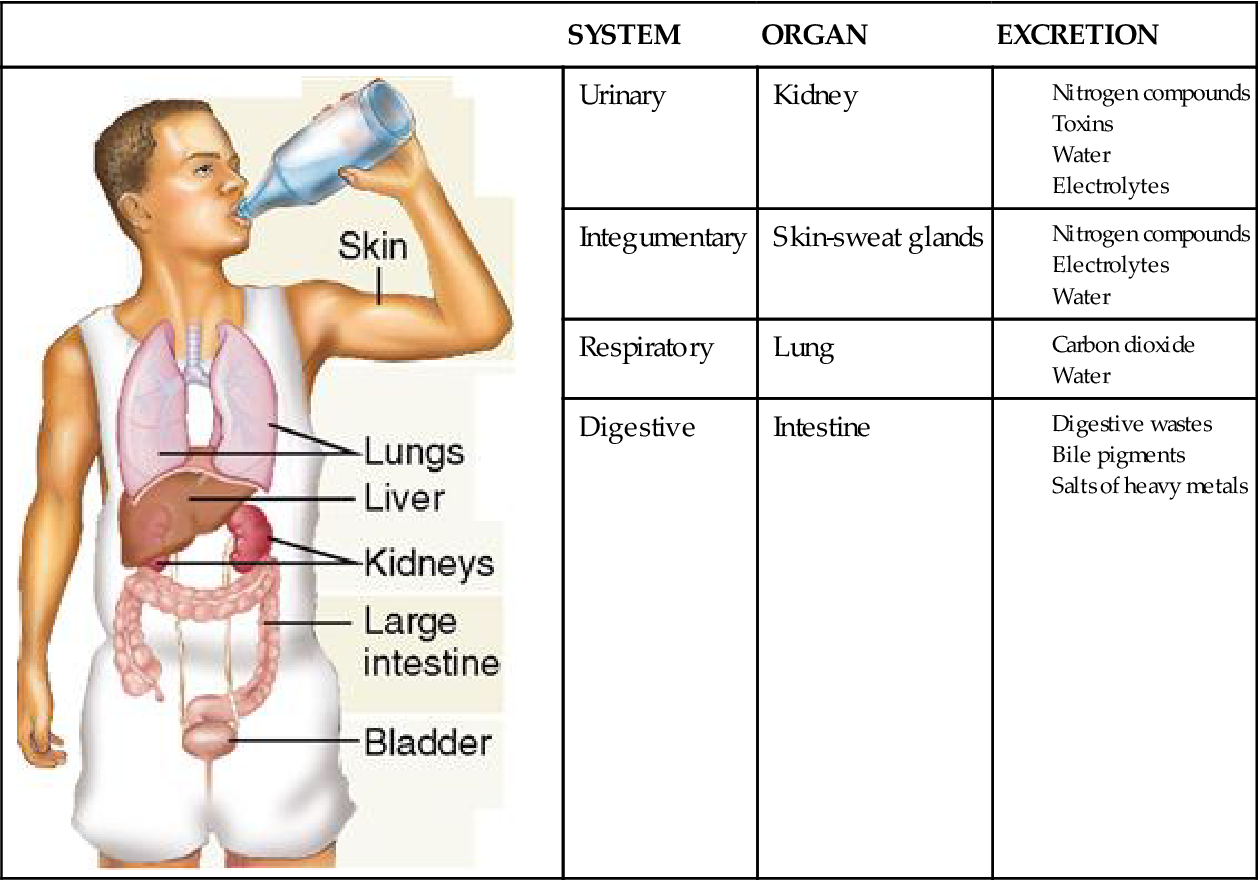
KIDNEY
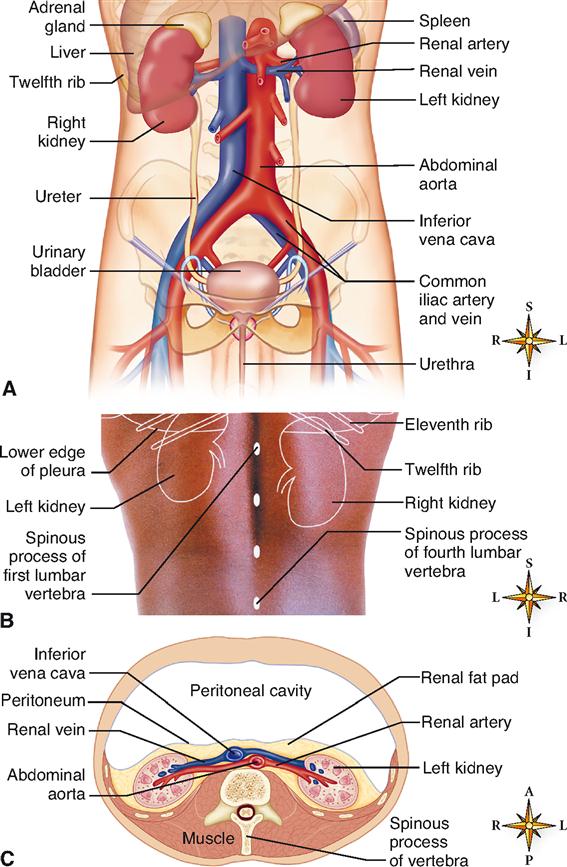
The kidneys resemble lima beans in shape, that is, roughly oval with a medial indentation (Figure 31-1, A). An average-sized kidney measures approximately 11 cm by 7 cm by 3 cm (4.3 by 2.7 by 1.2 inches). The left kidney is often slightly larger than the right. The kidneys lie in a retroperitoneal position, meaning posterior to the parietal peritoneum, against the posterior wall of the abdomen (Figure 31-1, C). They are located on either side of the vertebral column and extend from the level of the last thoracic vertebra (T12) to just above the third lumbar vertebra (L3). Note in Figure 31-1, B, that the superior or upper portions (poles) of both kidneys extend above the level of the twelfth rib and the lower edge of the thoracic parietal pleura. This anatomical relationship has important clinical implications (Box 31-2). Usually the right kidney is a little lower than the left, presumably because the liver takes up some of the space above the right kidney.
Box 31-2
 HEALTH matters
HEALTH mattersKidney Biopsy
Suspected disease of the kidney, such as renal cancer, often requires a needle biopsy to confirm the diagnosis. In these procedures a hollow needle is inserted through the skin surface and then guided into the diseased organ to withdraw a tissue sample for analysis. In a renal biopsy, tissue is removed from the lower rather than the upper or superior end of the diseased kidney. This avoids the possibility of the biopsy needle damaging the pleura and thereby causing a pneumothorax (see Box 27-3 on p. 835).
A heavy cushion of fat—the renal fat pad—normally encases each kidney and holds it in position. Connective tissue, the renal fasciae, anchors the kidneys to surrounding structures and also helps maintain their normal positions.
The medial surface of each kidney has a concave notch called the hilum. Renal blood vessels and other structures enter or leave the kidney through this notch. A tough, white fibrous capsule encases each kidney (Figure 31-2).
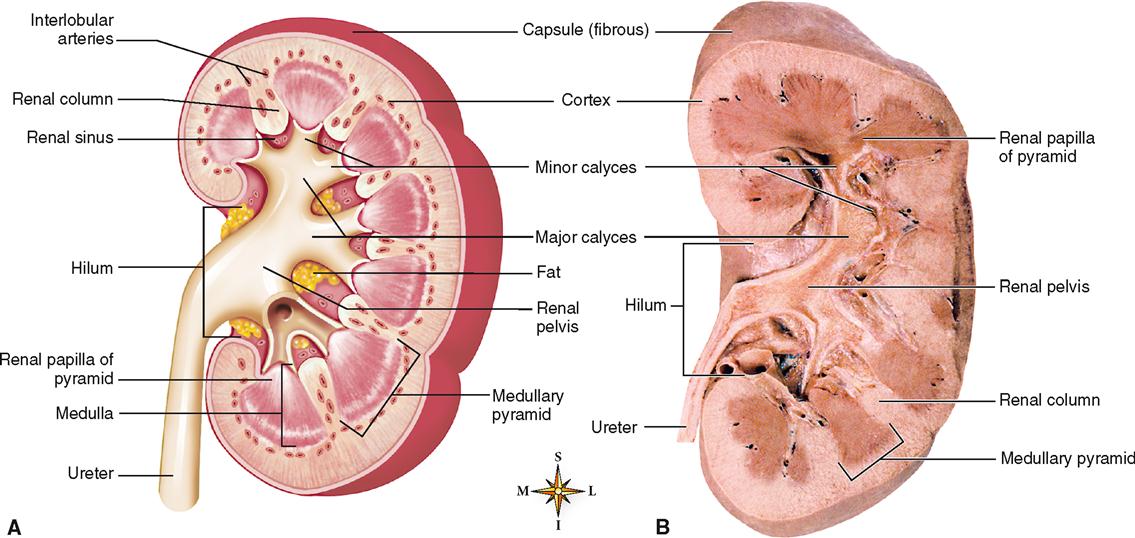
The coronal section of the right kidney shown in Figure 31-2 depicts the major internal structures of the kidney. Identify the renal cortex, or outer region, and the renal medulla, or inner region. A dozen or so distinct triangular wedges, the renal pyramids, make up much of the medullary tissue. The base of each pyramid faces outward, and the narrow papilla of each faces toward the hilum. Each renal papilla has multiple openings that release urine. Notice that the cortical tissue dips into the medulla between the pyramids, forming areas known as renal columns.
Each renal papilla (point of a pyramid) juts into a cuplike structure called a calyx. The calyces are considered the beginnings of the “plumbing system” of the urinary system, for it is here that urine leaving the renal papilla is collected for transport out of the body. The cups that drain the renal papillae directly are called minor calyces. These minor calyces are stemlike branches that join together to form larger branches called major calyces. The major calyces join together to form a large collection basin called the renal pelvis. The pelvis of the kidney narrows as it exits the hilum to become the ureter. Pelvis is Latin for “basin” and, like a lavatory basin, it collects fluid and quickly drains it away through a channel.
Blood Vessels of the Kidneys
The kidneys are highly vascular organs (Figure 31-3). Every minute about 1200 ml of blood flows through them. Stated another way, approximately one fifth of all the blood pumped by the heart per minute goes to the kidneys. From this fact one might guess, and correctly so, that the kidneys process the blood in important ways before returning it to the general circulation. A large branch of the abdominal aorta—the renal artery—brings blood into each kidney. As it nears the kidney, it divides into segmental arteries, which divide to become lobar arteries. Between the pyramids of the kidney’s medulla, the lobar arteries branch to form interlobar arteries that extend out toward the cortex, then arch over the bases of the pyramids to form the arcuate arteries. From the arcuate arteries, interlobular arteries penetrate the cortex. Because they radiate through the cortex, the interlobular arteries are sometimes called the cortical radiate arteries.
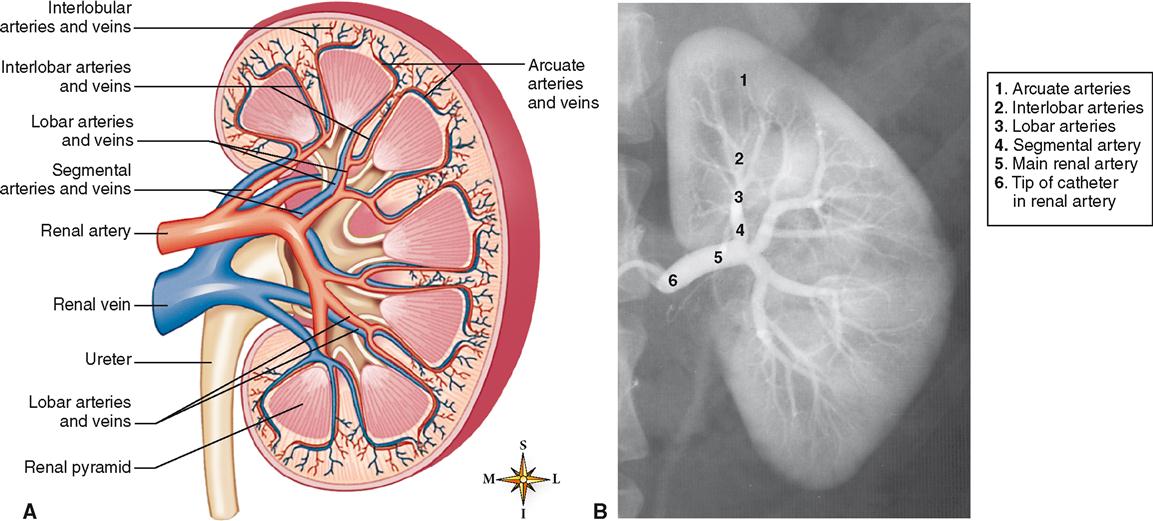
Branches of the interlobular arteries called afferent arterioles carry blood directly to the tiny functional units of the kidney called nephrons. We discuss the structure and function of nephrons later in this chapter, where we will resume outlining the path of blood flow—this time at a microscopic level.
Knowing the pathway of blood flow in the kidney is important for understanding how the kidney works. We continue the story later in the chapter. We put all the pieces together in an overview in Tracing Blood Flow in the Kidney online at A&P Connect.

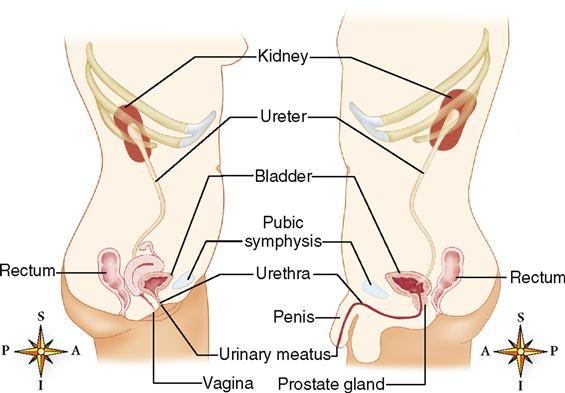
URETER
The ureters, about 28 to 34 cm in length, are the two tubes that actively convey urine from the kidneys to the urinary bladder (see Figure 31-1, A). They begin on each side at the narrow outlet of the renal pelvis on a level with the first lumbar vertebra (L1). Each ureter is retroperitoneal and courses into the pelvis until it reaches the bladder, where it attaches to the bottom of the bladder (Figure 31-4). It then runs at an angle for about 2 cm through the bladder wall and opens at the lateral angles of the trigone (floor) of the bladder (Figure 31-5). Because of its oblique course through the bladder wall, the ends of the tube close and act as valves when the bladder is full, thus preventing backflow of urine (Figure 31-6).
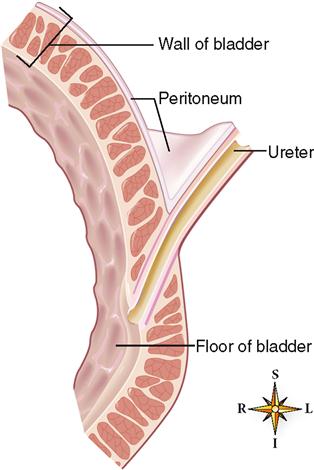
The ureter is lined with transitional epithelium, which permits stretching without damage to the epithelial lining. This feature permits either high or low rates of flow through the ureters.
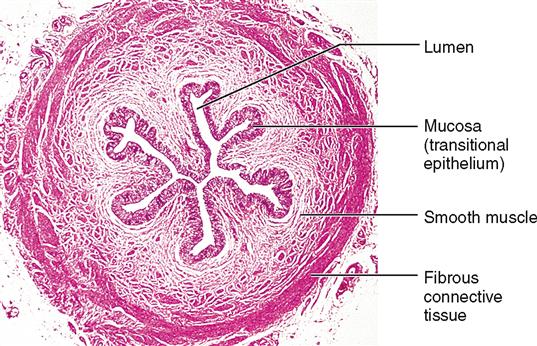
In females, the ureters are in close proximity to the ovaries and cervix of the uterus and, in males, in close proximity to the seminal vesicles and near the prostate gland (see Figure 31-4). Each ureter is composed of three layers of tissue: a mucous lining, a muscular middle layer, and a fibrous outer layer (Figure 31-7). The muscular layer is composed of smooth muscle, which propels the urine by peristalsis. The rate and strength of peristalsis increase with increasing urine volume.
URINARY BLADDER
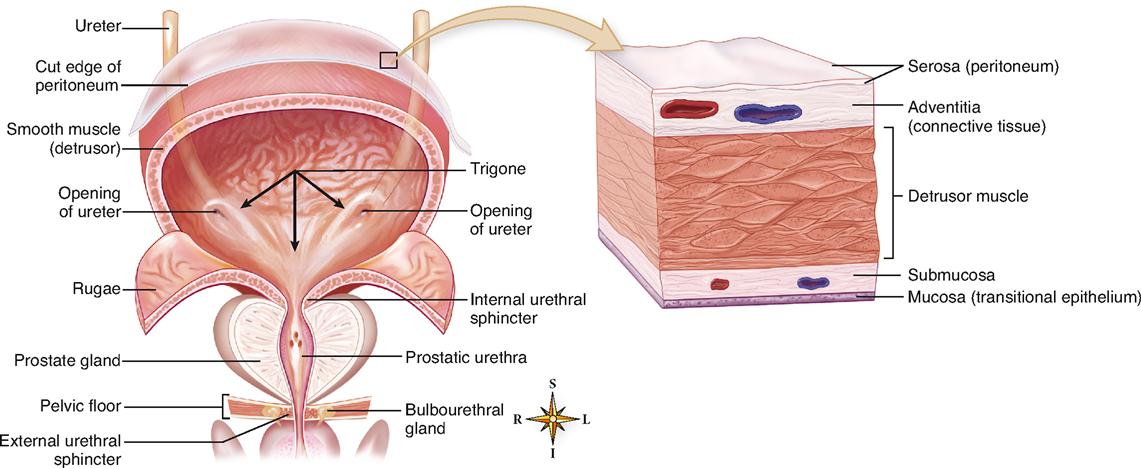
The urinary bladder is a muscular, collapsible bag that is located directly behind the pubic symphysis and in front of the rectum (see Figure 31-4). It lies below the parietal peritoneum, which covers only its superior surface (see Figure 31-5). The remainder of the bladder surface is covered by a fibrous adventitia. In women it sits on the anterior of the vagina and in front of the uterus, whereas in men, it rests on the prostate.
The wall of the bladder is made mostly of smooth muscle tissue (see Figure 31-5). Often called the detrusor muscle, the muscle layer is formed by a network of crisscrossing bundles of smooth muscle fibers. The bundles run in all directions: circular, oblique, and lengthwise. The bladder is lined with mucous transitional epithelium that forms folds called rugae (see Figure 31-5). Because of the folds and the extensibility of transitional epithelium, the bladder can distend considerably. There are three openings in the floor of the bladder—two from the ureters and one into the urethra. The ureter openings lie at the posterior corners of the triangle-shaped floor—the trigone—and the urethral opening lies at the anterior, lower corner.
The bladder performs two major functions:
URETHRA
The urethra is a small tube lined with mucous membrane (transitional epithelium) that leads from the floor of the bladder (trigone) to the exterior of the body. In females, it lies directly behind the pubic symphysis and anterior to the vagina as it passes through the muscular floor of the pelvis (Figure 31-8). It extends down and forward from the bladder for a distance of about 3 cm (1.2 inches) and ends at the external urinary meatus (see Figure 31-4). The functional importance of the relationship of the urethra and vagina to the muscular pelvic floor, especially after vaginal delivery of a baby, will be discussed in Chapter 35. The male urethra, on the other hand, extends along a winding path for about 20 cm (7.9 inches) (see Figure 31-4). The male urethra passes through the center of the prostate gland just after leaving the bladder. Within the prostate, it is joined by two ejaculatory ducts. After leaving the prostate, the urethra extends down, forward, then up to enter the base of the penis. It then travels through the center of the penis and ends as a urinary meatus at the tip of the penis.
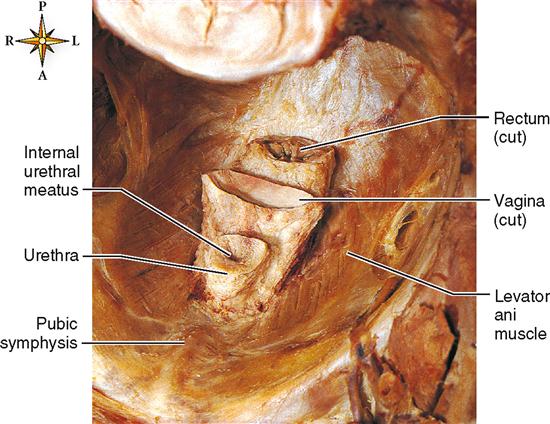
Because the male urethra is joined by the ejaculatory ducts, it serves as a pathway for semen (fluid containing sperm) as it is ejaculated out of the body through the penis. Thus we can say that the male urethra is a part of two different systems: the urinary system (when it is used to void urine) and the reproductive system (when it is used to ejaculate semen). Urine is prevented from mixing with semen during ejaculation by a reflex closure of sphincter muscles guarding the bladder’s opening. The female urethral tract is separate from the lower reproductive tract (vagina), which lies just behind the urethra (see Figure 31-8).
Micturition
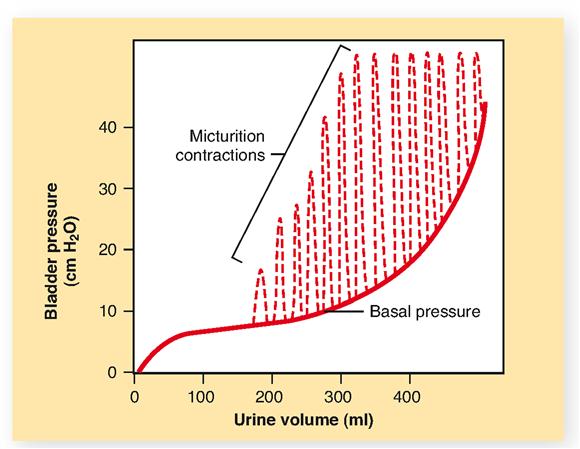
The mechanism for urinating begins with involuntary contractions of the detrusor muscle of the bladder wall (see Figure 31-5). Urination is also called voiding the bladder or micturition. Figure 31-9 shows that as the pressure of urine against the inside of the bladder wall increases with urine volume, involuntary micturition contractions develop. This rapid succession of involuntary contractions triggered by a parasympathetic reflex get stronger and stronger as the bladder fills and the urine volume and pressure increase. The parasympathetic reflex also causes the internal urethral sphincter muscles to relax at the same time. The internal urethral sphincters include a ringlike part of the detrusor muscle of the bladder wall, as you can see in Figure 31-5. The relaxation of these internal sphincters along with the micturition contractions of the bladder wall can force urine out of the bladder and through the urethra.
Thankfully, most people learn how to consciously regulate voluntary contraction of the external urethral sphincter muscles to stop voiding—that is, until a suitable time. Figure 31-5 shows that the skeletal muscles of the pelvic floor, including the levator ani muscle, act as external urethral sphincters (see also Figure 11-15 on p. 321). Voluntary control of micturition (voiding, or urination) is possible only if the nerves supplying the pelvic floor, the projection tracts of the central nervous system (CNS), and the motor areas of the brain are all intact and functioning properly. Learning this function is not possible until the nervous system matures sufficiently—making voluntary control of urination impossible during infancy and very early childhood.
Injury to any of these parts of the nervous system, by a cerebral hemorrhage or a spinal cord injury, for example, results in involuntary emptying of the bladder at intervals. Involuntary micturition is called incontinence (see Mechanisms of Disease, p. 994). In the average bladder, 250 ml of urine causes a moderately distended sensation and therefore the desire to void.
Microscopic Structure
Over a million microscopic functional units named nephrons make up the bulk of each kidney. The shape of the nephron is unusual, unmistakable, and uniquely suited to its function of blood plasma processing and urine formation (Figure 31-10). It resembles a tiny funnel with a long, winding stem about 3 cm (1.2 inches) long.
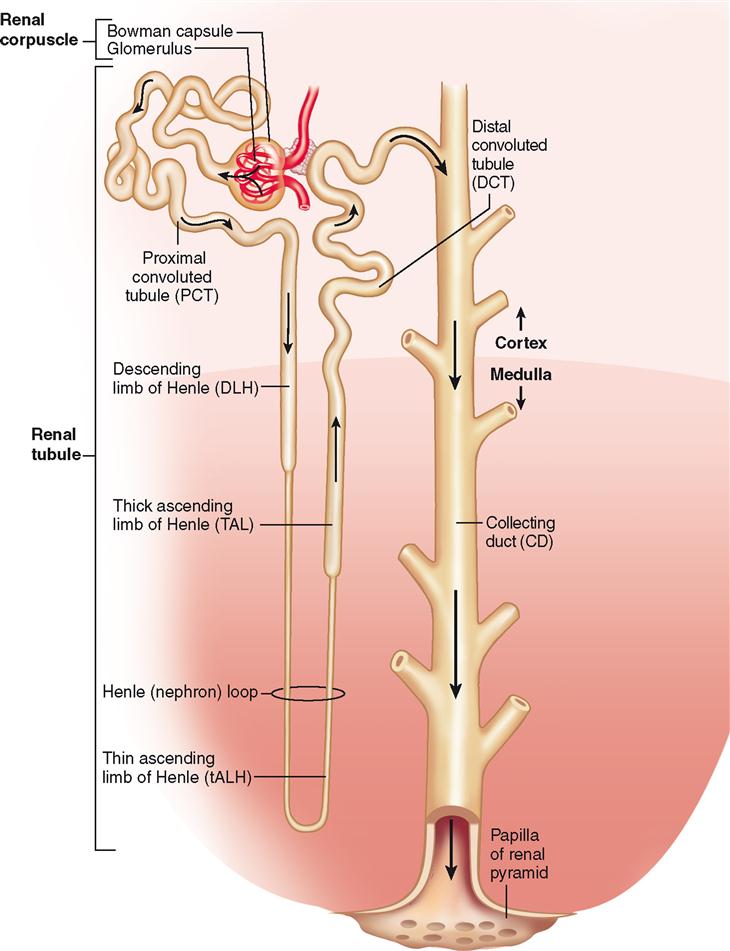
As Figure 31-10 shows, each nephron is made up of two main regions: the renal corpuscle and the renal tubule. Fluid is filtered out of the blood in the renal corpuscle, then the filtrate flows through the renal tubule and collecting duct—where much of the filtrate is returned to the blood. The remaining filtrate leaves the collecting duct as urine. We will come back to the processing of filtrate later. For now, we explore the structure of the nephron and collecting duct. Here are the main structures we explore, listed in the order in which fluid flows through them:


As you read the brief description of each of these microscopic structures, refer often to Figure 31-10, which shows a schematic diagram of a complete nephron.
For our purposes, the simplified scheme of microscopic renal anatomy shown in Figure 31-10 works well. However, some renal biologists prefer the more elaborate scheme sketched out in a Detailed Map of Nephron online at A&P Connect.

NEPHRON
Renal Corpuscle
The renal corpuscle is the first part of the nephron and is made up of the Bowman capsule and glomerulus. Formation of a renal corpuscle is sometimes compared to pushing your fist into the end of an inflated balloon. The mechanism is shown in Figure 31-11. Note that as the glomerular tuft of capillaries pushes into the balloon, it becomes surrounded by a double-walled cup with parietal (outer) and visceral (inner) walls—the Bowman capsule (Figure 31-12).
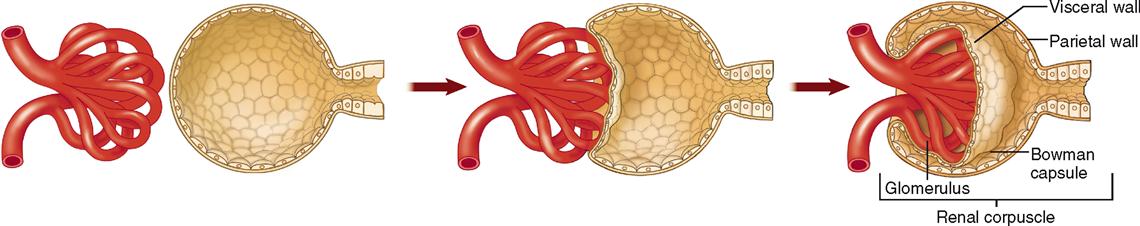
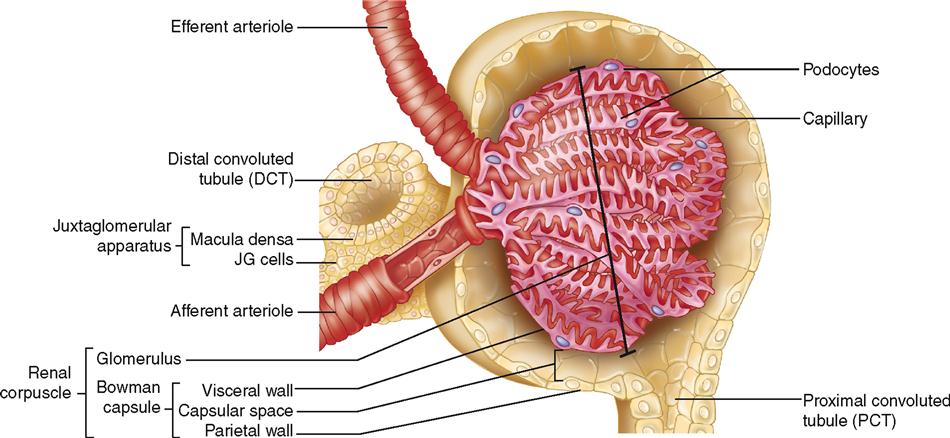
Fluid from the blood first filters out of the glomerulus and then into the Bowman capsule. We begin our discussion with the Bowman capsule first, however, because it will then make it easier to understand the structure and function of the glomerulus.
Bowman Capsule
The Bowman capsule is the cup-shaped mouth of a nephron. It is sometimes called the glomerular capsule. The capsule is formed by two layers of epithelial cells with a space, called the capsular space (Bowman space), between them (Figure 31-13). Fluids, waste products, and electrolytes that pass through the porous glomerular capillaries and enter this space constitute the filtrate, which will be processed in the nephron to form urine.
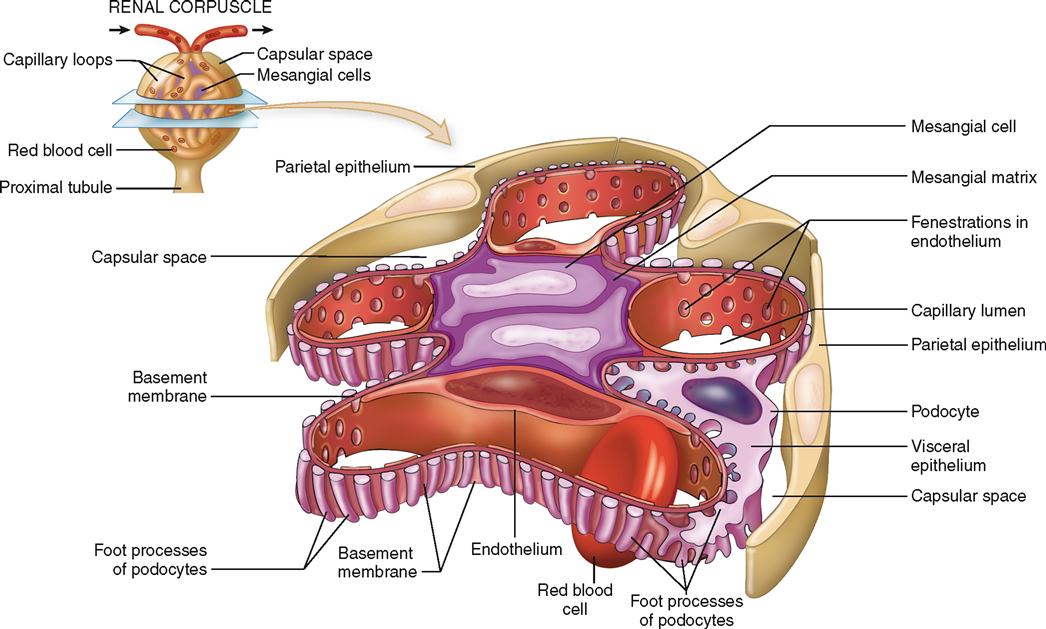
The parietal, or outer, wall is composed of simple squamous epithelium. It plays no role in the production of glomerular filtrate. The visceral (inner) wall, however, is quite different. It is composed of special epithelial cells called podocytes (meaning “cells with feet”). The scanning electron micrograph in Figure 31-14 reveals the odd shapes of podocytes. Notice that the primary branches extending from the cell bodies divide into a network of branches that terminate in little “feet” called pedicels. The pedicels are packed so closely together that only narrow slits of space lie between them. These spaces are called filtration slits. The slits are not merely open spaces, however. Within them is a mesh of fine connective tissue fibers called the slit diaphragm that prevents the slits from enlarging under pressure while still maintaining permeability of the slit. The slit diaphragm is an important component of the filter mechanism because it prevents many large macromolecules, such as proteins, from passing through.
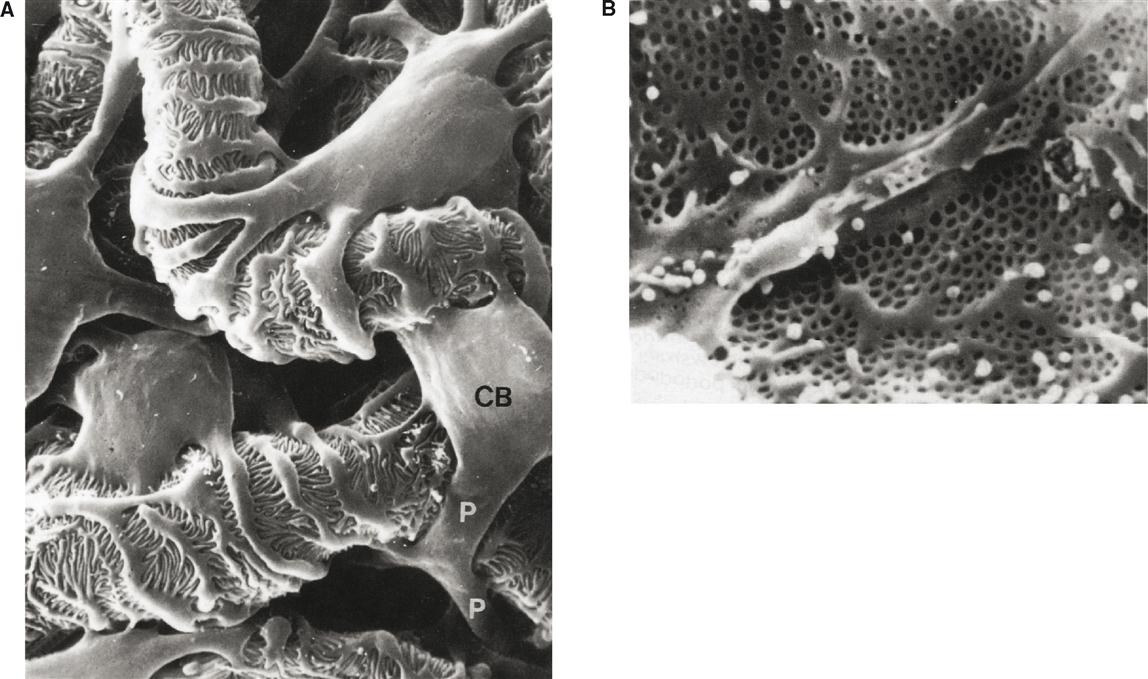
Glomerulus
The glomerulus is probably the body’s most well known capillary network and is surely one of its most important ones for survival. Its relationship to the Bowman capsule is clearly visible in Figures 31-11 and 31-12. Notice in both Figures that an afferent arteriole leads into the glomerular network and an efferent arteriole leads out.
Like all capillaries, glomerular capillaries have thin, membranous walls that are composed of a single layer of endothelial cells. Many pores, or fenestrations (meaning “windows”), are present in the glomerular endothelium (see Figure 31-14, B
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


