CHAPTER 8 The Special Senses
The evolution of vertebrates shows a trend called cephalization in which special sensory organs develop in the heads of animals, along with the corresponding development of the brain. These special sensory systems, which include the visual, auditory, vestibular, olfactory, and gustatory systems, detect and analyze light, sound, and chemical signals in the environment, as well as signal the position and movement of the head. The stimuli detected and transduced by these systems are most familiar to us when they provide conscious awareness of our environment, but they are equally important as sensory input for reflexive and subconscious behavior.
THE VISUAL SYSTEM
Structure of the Eye
The wall of the eye is composed of three concentric layers (Fig. 8-1). The outer layer, or the fibrous coat, includes the transparent cornea, with its epithelium, and the opaque sclera. The middle layer, or vascular coat, includes the iris and the choroid. The iris contains both radially and circularly oriented smooth muscle fibers, which make up the pupillary dilator and sphincter muscles. The choroid is rich in blood vessels that support the outer layers of the retina, and it also contains pigment. The innermost layer of the eye is the retina, which is embryologically derived from the diencephalon and is therefore part of the central nervous system (CNS). The functional part of the retina covers the entire posterior aspect of the eye except for the optic nerve head or optic disc, which is where the optic nerve axons leave the retina. Because there are no receptors at this location, it is often referred to as the anatomic “blind spot” (Fig. 8-1).
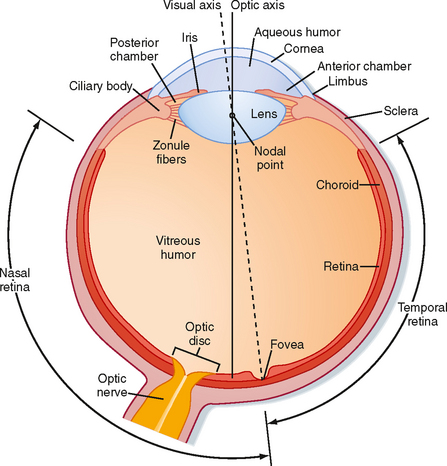
Figure 8-1 View of a horizontal section of the right eye.
(Redrawn from Wall GL: The Vertebrate Eye and Its Adaptive Radiation. Bloomfield Hills, MI, Cranbrook Institute of Science, 1942.)
A number of functions of the eyes are under muscular control. Externally attached extraocular muscles aim the eyes toward an appropriate visual target (see Chapter 9). These muscles are innervated by the oculomotor (cranial nerve [CN] III), trochlear (CN IV), and abducens (CN VI) nerves. Several muscles are also found within the eye (intraocular muscles). The muscles in the ciliary body control lens shape and thereby the focus of images on the retina. The pupillary dilator and sphincter muscles allow the iris to control the amount of light entering the eye, similar to the diaphragm of a camera. The dilator is activated by the sympathetic nervous system, whereas the sphincter and ciliary muscles are controlled by the parasympathetic nervous system (through the oculomotor nerve) (see Chapter 11).
Light enters the eye through the cornea and passes through a series of transparent fluids and structures that are collectively called the dioptric media. These fluids and structures consist of the cornea, aqueous humor, lens, and vitreous humor. The aqueous humor, located in the anterior and posterior chambers and the vitreous humor in the space behind the lens, respectively, help maintain the shape of the eye. The aqueous humor is secreted by the epithelium of the ciliary body into the posterior chamber of the eye. It then circulates through the pupil and into the anterior chamber, where it is drained into the venous system by the canal of Schlemm. Aqueous humor pressure, which is normally less than 22 mm Hg, determines the pressure within the eye. The vitreous humor is a gel composed of extracellular fluid that contains collagen and hyaluronic acid; unlike aqueous humor, however, it turns over very slowly.
Normally, light from a visual target is focused sharply on the retina by the cornea and lens, which bend or refract the light. The cornea is the major refractive element of the eye, with a refractive power of 43 diopters* (D). However, unlike the cornea, the lens can change shape and vary its refractive power between 13 and 26 D. Thus, the lens is responsible for adjusting the optical focus of the eye. Suspensory ligaments (or zonule fibers), which attach to the wall of the eye at the ciliary body (Fig. 8-1), hold the lens in place. When the muscles in the ciliary body are relaxed, the tension exerted by the suspensory ligaments flattens the lens. When the ciliary muscles contract, the tension on the suspensory ligaments is reduced; this process allows the somewhat elastic lens to assume a more spherical shape. The ciliary muscles are activated by the parasympathetic nervous system (via the oculomotor nerve).
Although the optic axis of the human eye passes through the nodal point of the lens and reaches the retina at a point between the fovea and the optic disc (Fig. 8-1), the eye is directed by the oculomotor system to a point, called the fixation point, on the visual target. Light from the fixation point passes along the optic axis, and is focused on the fovea. Light from the remainder of the visual target falls on the retina surrounding the fovea.
Retina
Layers of the Retina
The 10 layers of the retina are shown in Figure 8-2. The retina begins with the pigmented epithelium (layer 1), which is just inside the choroid. The pigment cells have tentacle-like processes that extend into the photoreceptor layer (layer 2) and surround the outer segments of the rods and cones. These processes prevent transverse scatter of light between photoreceptors. In addition, they serve a mechanical function in maintaining contact between layers 1 and 2 so that the pigmented epithelium can (1) provide nutrients and remove waste from the photoreceptors; (2) phagocytose the ends of the outer segments of the rods, which are continuously shed; and (3) reconvert metabolized photopigment into a form that can be reused after it is transported back to the photoreceptors.
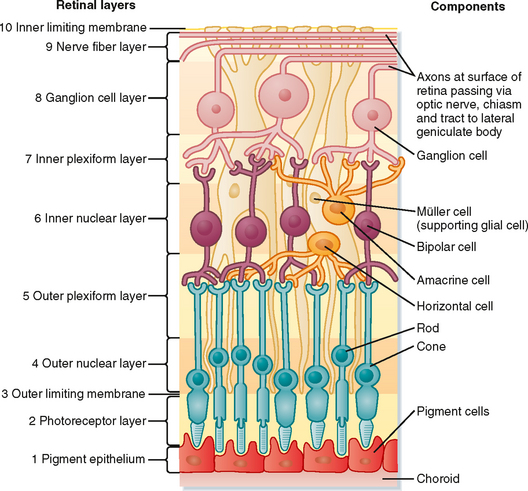
Light rays that originate from different parts of the visual target map onto the photoreceptor array of layer 2 in a point-to-point fashion. Retinal glial cells, known as Müller cells, play an important role in maintaining the internal geometry of the retina. Müller cells are oriented radially, parallel to the light path through the retina. The outer ends of Müller cells form tight junctions with the inner segments of the photoreceptors. The numerous connections made between Müller cells and the inner segments give the appearance of a continuous layer, the outer limiting membrane (layer 3 of the retina).
Structure of Photoreceptors: Rods and Cones
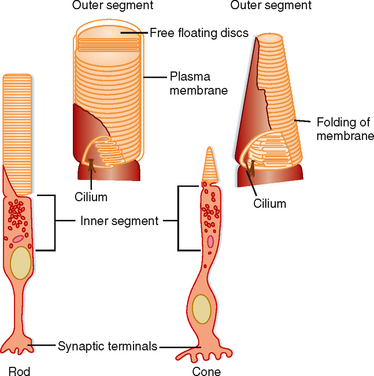
Each rod or cone photoreceptor cell is composed of a cell body (in layer 4), an inner segment and an outer segment that extend into layer 2, and a synaptic terminal that projects into layer 5 (Fig. 8-3). The outer segments of cones are not as long as those of rods, and they contain stacks of disc membranes formed by infoldings of the plasma membrane. The outer segments of rods are longer, and the stacks of membrane discs float freely in the outer segment after having disconnected from the plasma membrane when formed at the base. Both sets of discs are rich in photopigment molecules, but the greater photopigment density of rods partly accounts for their greater sensitivity to light. A single photon can elicit a rod response, whereas several hundred photons may be required for a cone response.
Regional Variations in the Retina
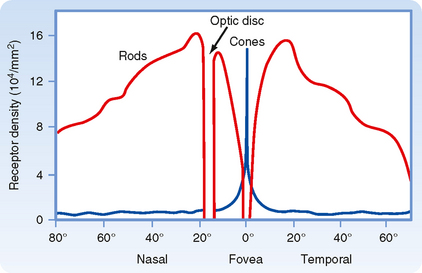
The fovea, which is a depression in the macula lutea, is the region of the retina that has the very highest visual resolution. Correspondingly, the image from the fixation point is focused on the fovea. The retinal layers in the foveal region are unusual because several of them appear to be pushed aside into the surrounding macula. Because light can reach the foveal photoreceptors without having to pass through the inner layers of the retina, both image distortion and light loss are minimized. The fovea has cones with unusually long and thin outer segments. This cone shape permits high packing density. In fact, cone density is maximal in the fovea, and this high density provides for high visual resolution, as well as high quality of the image (Fig. 8-4).
Visual Transduction
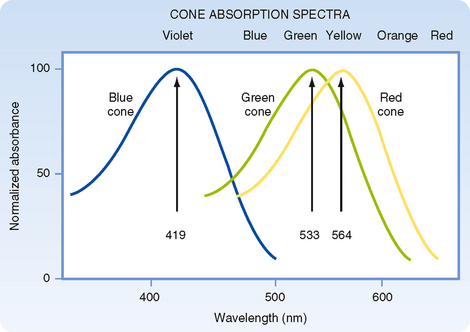
Light energy must be absorbed for it to be detected by the retina. Light absorption is accomplished by the visual pigments, which are located in the outer segments of the rods and cones. The pigment found in the outer segments of rods is rhodopsin, or visual purple (so named because it has a purple appearance after green or blue light have been absorbed), and it absorbs light best at a wavelength of 500 nm. Three variants of visual pigment are found in cones, and these cone pigments absorb best at 419 nm (blue), 533 nm (green), or 564 nm (red). However, the absorption spectrum of visual pigments is broad so that they overlap considerably (Fig. 8-5).
Rhodopsin is formed when a retinal isomer, 11-cis retinal, is combined with a glycoprotein known as opsin. When rhodopsin absorbs light, it is “boosted” to a higher energy state. This boost causes a series of chemical changes that lead to isomerization of 11-cis retinal to all-trans retinal, release of the bond with opsin, and conversion of retinal to retinol. Separation of all-trans retinal from opsin causes bleaching of the visual pigment; that is, the pigment loses its purple color.
In darkness, photoreceptors are slightly depolarized (around −40 mV) because cGMP-gated Na+ channels (Fig. 8-6, A) in their outer segments are open, thereby increasing gNa and driving the membrane potential toward the Na+ equilibrium potential. This net influx of Na+ results in a continuous current, called the dark current. As a consequence of this constant depolarization, the neurotransmitter glutamate is tonically released at the rod cell’s synapses. Intracellular [Na+] is kept at a steady-state level by the pumping action of Na+,K+-ATPase.
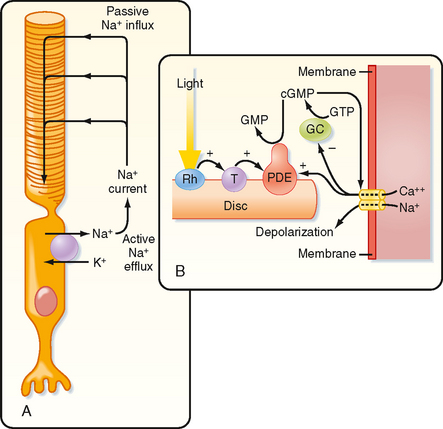
When light is absorbed, the photoisomerization of rhodopsin activates a G protein called transducin (Fig. 8-6, B). This G protein, in turn, activates cyclic guanosine monophosphate phosphodiesterase, which is associated with the rhodopsin-containing discs, hydrolyzes cGMP to 5′-GMP, and lowers the cGMP concentration in the rod cytoplasm. The reduction in cGMP leads to closing of the cGMP-gated Na+ channels, hyperpolarization of the photoreceptor membrane, and a reduction in the release of transmitter. Thus, cGMP acts as a “second messenger” to translate reception of a photon by the photopigment into a change in membrane potential.
The extraordinary sensitivity of rods, which can signal the capture of a single photon, is enhanced by an amplification mechanism such that photoactivation of only one rhodopsin molecule can activate hundreds of transducin molecules. In addition, each phosphodiesterase molecule hydrolyzes thousands of cGMP molecules per second. Similar events occur in cones, but the membrane hyperpolarization occurs much more quickly than in rods, and requires thousands of photons.
Color Vision
The three visual pigments in the cone outer segments have opsins that differ from the opsin found in rhodopsin. As a result of these differences, the three types of cone pigments absorb light best at different wavelengths. Although the cone pigments have maximum efficiency closer to violet, green, and yellow wavelengths, they are referred to as blue, green, and red pigments, respectively (Fig. 8-5).
According to the trichromacy theory, these differences in absorption efficiency are presumed to account for color vision because a suitable mixture of three colors can produce any other color. However, a neural system must also exist for the analysis of color brightness because the amount of light absorbed by a visual pigment, as well as the subsequent response of the cell, depends on both the wavelength and the intensity of the light (Fig. 8-5). Two or three of the cone pigments may absorb a particular wavelength of light, but the amount absorbed by each will differ according to their efficiencies at that wavelength. If the intensity of the light is increased (or decreased), all will absorb more (or less), but the ratio of absorption among them will remain constant. Consequently, there must be a neural mechanism to compare the absorption of light of different wavelengths by the different types of cones for the visual system to distinguish different colors. At least two different kinds of cones are required for color vision. The presence of three kinds decreases the ambiguity in distinguishing colors when all three absorb light, and it ensures that at least two types of cones will absorb most wavelengths of visible light.
Retinal Circuitry
A diagram of the basic circuitry of the retina is shown in Figure 8-7. Photoreceptors (R) synapse on the dendrites of bipolar cells (B) and horizontal cells (H) in the outer plexiform layer. The horizontal cells make reciprocal synaptic connections with photoreceptor cells, are electrically coupled to other horizontal cells, and receive input from interplexiform cells (I). Bipolar cells synapse on the dendrites of ganglion cells (G) and on the processes of amacrine cells (A) in the inner plexiform layer. Amacrine cells connect with ganglion cells, other amacrine cells, and interplexiform cells.
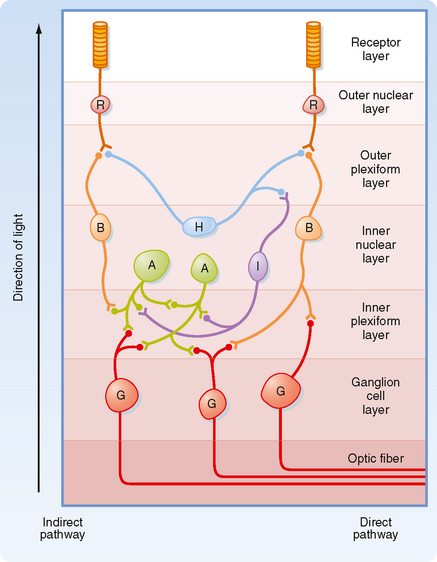
Several features of this circuitry are noteworthy. Input to the retina is provided by light striking the photoreceptors. The output is carried by axons of the retinal ganglion cells to the brain. Information is processed within the retina by the interneurons. The most direct pathway through the retina is from a photoreceptor to a bipolar cell and then to a ganglion cell (Fig. 8-7). More indirect pathways that provide for intraretinal signal processing involve photoreceptors, bipolar cells, amacrine cells, and ganglion cells, as well as horizontal cells to provide lateral interactions between adjacent pathways. Interplexiform cells allow interactions to occur from the inner to the outer retina.
Contrasts in Rod and Cone Pathway Functions
Cones have a higher threshold to light and thus are not activated in dim light after dark adaptation. However, they operate very well in daylight. They provide high-resolution vision because only a few cones converge onto individual bipolar cells in the cone pathways. Moreover, no convergence occurs in the fovea where the cones make one-to-one connections to bipolar cells. As a result of the reduced convergence, cone pathways have very small receptive fields and can resolve stimuli that originate from sources very close to each other. Cones also respond to sequential stimuli with good temporal resolution. Finally, cones have three different cone photopigments. Thus, they can discriminate relative spectral content independent of absolute intensity and therefore provide for color vision. Loss of cone function results in functional blindness; rod vision is not sufficient for normal visual requirements.
Receptive Field Organization
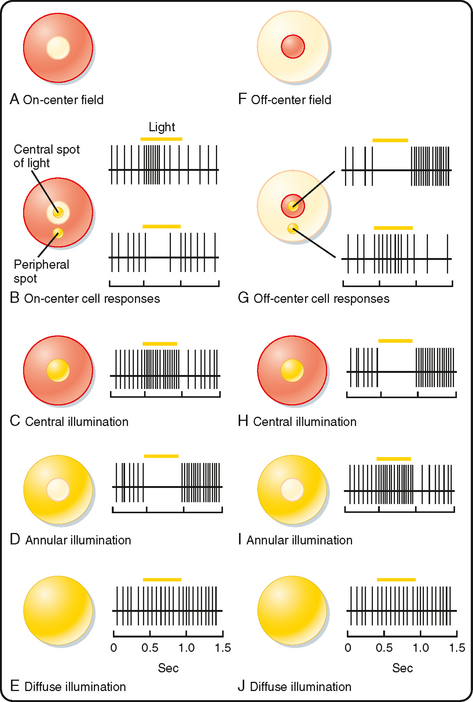
A bipolar cell that receives input from a photoreceptor can have either of two types of receptive fields, as shown in Figure 8-8. Both are described as having a center-surround organization in which the light that strikes the central region of the receptive field either excites or inhibits the cell, whereas the light that strikes the annular region that surrounds the central portion has the converse effect. The receptive field with a centrally located excitatory region surrounded by an inhibitory annulus is called an on-center, offsurround receptive field (Fig. 8-8, A). Bipolar cells with such a receptive field are described as “on” bipolars. The other type of receptive field has an off-center, on-surround arrangement, which characterizes “off” bipolars (Fig. 8-8, F).
(Redrawn from Squire LR et al [eds]: Fundamental Neuroscience. San Diego, CA, Academic Press, 2002.)
The central property of bipolar cell receptive fields is due to only a few directly connected photoreceptors. The antagonistic surround response is due to light impinging on adjacent photoreceptors, which changes the activity of horizontal cells. This pathway through the horizontal cells results in a response that is opposite in sign to that produced directly by the photoreceptors that mediate the center response. The basis for this is that horizontal cells, like “off” bipolars, are hyperpolarized in the light and, because they are electrically coupled to each other by gap junctions, have very large receptive fields. Darkness in the periphery of a bipolar cell’s receptive field (such as an annulus that does not affect the photoreceptors to which it is directly connected) will depolarize neighboring photoreceptors and horizontal cells. Depolarized horizontal cells release GABA onto central (and peripheral) photoreceptor terminals, reducing their release of glutamate. Thus, when a darkness surrounds central illumination, there is increased excitation of on-center bipolars. There is a complementary effect on off-center bipolars when a bright annulus surrounds a central dark spot (Fig. 8-8).
Ganglion cells may receive dominant input from bipolar cells, dominant input from amacrine cells, or mixed input from amacrine and bipolar cells. When amacrine cell input dominates, the receptive fields of ganglion cells tend to be diffuse, and they are either excitatory or inhibitory. Most ganglion cells are dominated by bipolar cell input and have a center-surround organization, similar to that of bipolar cells (Fig. 8-8).
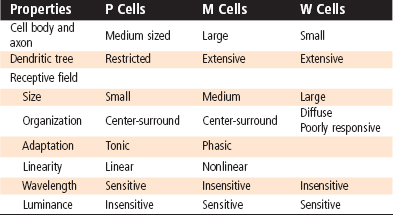
P, M, and W Cells
Several of the physiological differences among these cell types correspond to morphological differences (Table 8-1). For example, P cells have small receptive fields (which corresponds to smaller dendritic trees) and more slowly conducting axons than M cells do. In addition, P cells show a linear response in their receptive field; that is, they respond with a sustained, tonic discharge of action potentials to maintained light but do not signal shifts in the pattern of illumination as long as the overall level of illumination is constant. Thus, a small object entering a P cell’s central receptive field will change its firing, but continued movement within the field will not be signaled. P cells respond differently to different wavelengths of light. Because there are blue, green, and red cones, many combinations of color properties are possible, but in fact P cells have been shown to have only opposing responses to red and green or to blue and yellow (a combination of red and green). They may have center-surround antagonism in which one color excites the center while the other inhibits the surround (or vice versa), or one color might excite the entire receptive field while another inhibits it (e.g., R+G− describes a cell that is excited by red and inhibited by green). These mechanisms can greatly reduce the ambiguity of color detection caused by the overlap in cone color sensitivity and may provide a substrate for the opponency process observations.
The Visual Pathway
Retinal ganglion cells transmit information to the brain by way of the optic nerve, optic chiasm, and optic tract. Figure 8-9 shows the relationships between a visual target (arrow), the retinal images of the target in the two eyes, and the projections of retinal ganglion cells to the two hemispheres of the brain. The eyes and the optic nerves, chiasm, and tract are viewed from above.
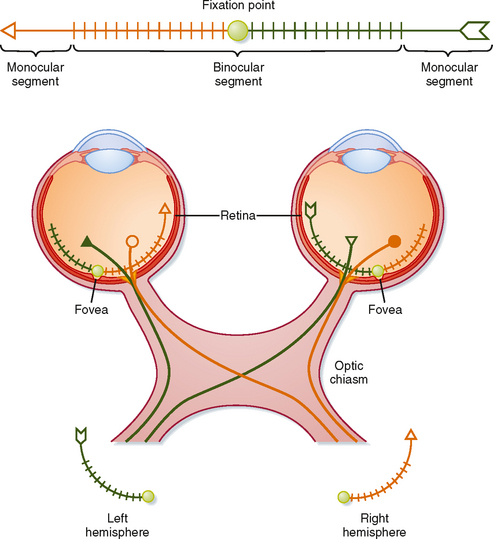
The visual target, an arrow, is in the visual fields of both eyes (Fig. 8-9) and, in this case, is so long that it extends into the monocular segments of each retina (i.e., one end of the target can be seen only by one eye and the other end only by the other eye). The shaded circle at the center of the target shows the fixation point. The image of the target is reversed on the retinas by the lens system. The left half of the visual target is imaged on the nasal retina of the left eye and the temporal retina of the right eye. Thus, the left visual field is seen by the left nasal retina and the right temporal retina. Similarly, the right half of the visual target is imaged on and seen by the left temporal retina and the right nasal retina. There is also an inversion in the vertical axis, with the upper visual field imaged on the lower retina and vice versa.
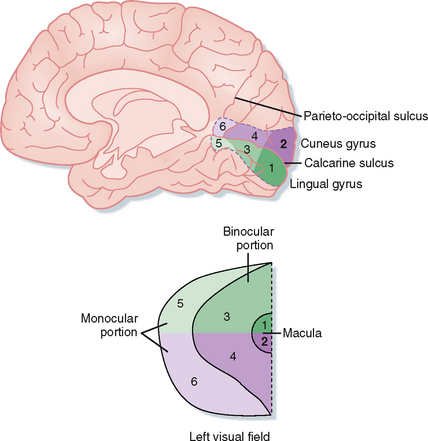
The projections of retinal ganglion cells may be uncrossed or crossed, depending on the location of the ganglion cell in the retina (Fig. 8-9). Axons from the temporal portion of each retina pass through the optic nerve, the lateral side of the optic chiasm, and the ipsilateral optic tract and terminate ipsilaterally in the brain. Axons from the nasal portion of each retina pass through the optic nerve, cross to the opposite side in the optic chiasm, and then pass through the contralateral optic tract to end in the contralateral side of the brain. This arrangement results in the representation of objects in the left field of vision in the right side of the brain and those in the right field of vision in the left side of the brain (Fig. 8-10).
< div class='tao-gold-member'>
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


