Objectives
The reader uses the knowledge he or she gained from the preceding chapters to predict the response of the respiratory system to 4 physiologic stresses: exercise, ascent to altitude, diving, and sleep.
- Identifies the physiologic stresses involved in exercise.
- Predicts the responses of the respiratory system to acute exercise.
- Describes the effects of long-term exercise programs (training) on the respiratory system.
- Identifies the physiologic stresses involved in the ascent to altitude.
- Predicts the initial responses of the respiratory system to the ascent to altitude.
- Describes the acclimatization of the cardiovascular and respiratory systems to residence at high altitudes.
- Identifies the physiologic stresses involved in diving.
- Predicts the responses of the respiratory system to various types of diving.
- Identifies the physiologic stresses involved in sleep.
- Relates the alterations that occur in the respiratory system during sleep to the pathophysiology of obstructive sleep apnea.
The Respiratory System under Stress: Introduction
This chapter is mainly intended to be a review of the preceding chapters of the book. The responses of the respiratory system to 4 physiologic stresses are examined as they relate to the material already covered; the discussions of the responses to each stress will therefore be brief and rather superficial. For a more complete discussion of each stress, consult the Suggested Readings at the end of this chapter.
Exercise and the Respiratory System
 Exercise increases the metabolism of the working muscles. It stresses the respiratory system by increasing the demand for oxygen and increasing the production of carbon dioxide. Moderate to strenuous levels of exercise also cause increased lactic acid production. The respiratory and cardiovascular systems must increase the volume of oxygen supplied to the exercising tissues and increase the removal of carbon dioxide and hydrogen ions from the body.
Exercise increases the metabolism of the working muscles. It stresses the respiratory system by increasing the demand for oxygen and increasing the production of carbon dioxide. Moderate to strenuous levels of exercise also cause increased lactic acid production. The respiratory and cardiovascular systems must increase the volume of oxygen supplied to the exercising tissues and increase the removal of carbon dioxide and hydrogen ions from the body.
The effects of exercise in an untrained person are mainly a function of an increase in the cardiac output coupled with an increase in alveolar ventilation.
As discussed at the end of Chapter 9, both the tidal volume and the breathing frequency are increased during exercise. The causes of the increased alveolar ventilation during exercise were discussed in that section.
The work of breathing is increased during exercise. Larger tidal volumes result in increased work necessary to overcome the elastic recoil of the lungs and chest wall during inspiration because the lungs are less compliant at higher lung volumes and because the elastic recoil of the chest wall is inward at high thoracic volumes. Of course, the greater elastic recoil tends to make expiration easier, but this is offset by other factors. The high airflow rates generated during exercise result in a much greater airways resistance component of the work of breathing. Greater turbulence and dynamic compression of airways secondary to active expiration combine to greatly increase the work of breathing. (Recall that during turbulent airflow 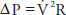 .) Increasing airflow rates especially increases the resistive work of breathing through the nose: Minute ventilations above about 40 L/min are normally accomplished by breathing through the mouth.
.) Increasing airflow rates especially increases the resistive work of breathing through the nose: Minute ventilations above about 40 L/min are normally accomplished by breathing through the mouth.
In normal adults, the resting minute ventilation (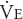 ) of 5 to 6 L/min can be increased to as much as 150 L/min during short periods of maximal exercise. Maximal increases in cardiac output during exercise are only in the range of 4 to 6 times the resting level in healthy adults compared with this 25-fold potential increase in minute ventilation. Therefore, it is the cardiovascular system rather than the respiratory system that is the limiting factor in exercise by healthy people.
) of 5 to 6 L/min can be increased to as much as 150 L/min during short periods of maximal exercise. Maximal increases in cardiac output during exercise are only in the range of 4 to 6 times the resting level in healthy adults compared with this 25-fold potential increase in minute ventilation. Therefore, it is the cardiovascular system rather than the respiratory system that is the limiting factor in exercise by healthy people.
As discussed in Chapter 9, at less strenuous levels of exercise, the increase in ventilation is accomplished mainly by increasing the tidal volume. During strenuous exercise, the tidal volume usually increases to a maximum of about 50% to 60% of the vital capacity of a normal subject, or about 2.5 to 3.0 L in an average-sized man. This increase in tidal volume appears to occur mainly at the expense of the inspiratory reserve volume, with the expiratory reserve volume somewhat less affected. An increase in the central blood volume (caused by increased venous return) may decrease the total lung capacity slightly. The residual volume and functional residual capacity may be unchanged or slightly elevated. The vital capacity may be slightly decreased or unchanged. The breathing frequency may increase to 40 to 50 breaths/min in healthy adults (and as high as 70 breaths per minute in children) with strenuous exercise.
The anatomic dead space may increase slightly in inspiration during exercise because of airway distention at high lung volumes; any alveolar dead space present at rest normally decreases as cardiac output increases. As a result, there is little change in physiologic dead space during exercise. Because the tidal volume does increase, however, the ratio of physiologic dead space to tidal volume (VD/VT) decreases.
The arterial 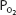 stays relatively constant during even strenuous exercise. Arterial
stays relatively constant during even strenuous exercise. Arterial 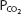 also stays relatively constant until anaerobic metabolism results in appreciable lactic acid generation. The hydrogen ions liberated directly stimulate alveolar ventilation and may cause arterial
also stays relatively constant until anaerobic metabolism results in appreciable lactic acid generation. The hydrogen ions liberated directly stimulate alveolar ventilation and may cause arterial 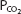 to fall a few millimeters of mercury below the resting arterial
to fall a few millimeters of mercury below the resting arterial 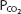 .
.
The regional differences in alveolar ventilation seen in upright lungs (which were discussed in Chapter 3) are probably slightly attenuated during exercise. The larger tidal volumes, occurring at the expense of both the inspiratory and expiratory reserve volumes, indicate that alveoli in more dependent regions of the lung are more fully inflated. However, these alveoli may also suffer airway collapse during active expirations. Similarly, alveoli in upper portions of the lung (with respect to gravity) should deflate more fully during expiration, resulting in greater ventilation of upper parts of the lung.
As already mentioned, the cardiac output increases linearly with oxygen consumption during exercise. This normally occurs more as a result of an autonomically mediated increase in heart rate than from an increase in stroke volume. An increased venous return, due to deeper inspiratory efforts (in addition to extravascular compression by the exercising muscles and by a decrease in venous capacitance), also contributes to the increase in cardiac output. Mean pulmonary artery and mean left atrial pressures increase, but the increase is not as great as the increase in pulmonary blood flow. This indicates a decrease in pulmonary vascular resistance. As discussed in Chapter 4, this decrease occurs passively by recruitment and distention of pulmonary vessels. Much of the recruitment of pulmonary blood vessels occurs in upper regions of the lung, thus tending to decrease the regional inhomogeneity of pulmonary blood flow discussed in Chapter 4. The expected effect of the deeper tidal volumes and active expirations that occur during exercise is to increase pulmonary vascular resistance. During active expiration the extraalveolar vessels should be compressed; during inspiration the alveolar vessels should be stretched. Mean pulmonary vascular resistance decreases, so the effects of recruitment and distention must be greater than the effects of stretching vessels and extravascular compression.
The more uniform regional perfusion that occurs during exercise results in a much more uniform matching of ventilation and perfusion throughout the lung. Studies done on normal subjects engaged in exercise in the upright position have demonstrated a greatly increased perfusion of upper regions of the lung, resulting in improved matching of ventilation and perfusion. Because in moderate to severe exercise ventilation increases more than perfusion, the whole lung ventilation-per-fusion ratio (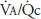 ) increases to a range of 2.0 to 4.0, as seen in Figure 11–1, where the
) increases to a range of 2.0 to 4.0, as seen in Figure 11–1, where the 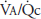 for the whole lung is approximately 2.5. A comparison of these findings with the ventilation-perfusion ratios seen at rest in the figure and in Figure 5–7 demonstrates the reduced “scatter” of ventilation-perfusion ratios during exercise. Thus, the location of the perfusion is better matched to the location of the ventilation during exercise, but the ventilation-perfusion ratios increase in most alveolar-capillary units. The increased ventilation-perfusion ratios may enhance the alveolar-capillary diffusion gradients for oxygen and carbon dioxide in many units. On the other hand, several investigations have demonstrated increased
for the whole lung is approximately 2.5. A comparison of these findings with the ventilation-perfusion ratios seen at rest in the figure and in Figure 5–7 demonstrates the reduced “scatter” of ventilation-perfusion ratios during exercise. Thus, the location of the perfusion is better matched to the location of the ventilation during exercise, but the ventilation-perfusion ratios increase in most alveolar-capillary units. The increased ventilation-perfusion ratios may enhance the alveolar-capillary diffusion gradients for oxygen and carbon dioxide in many units. On the other hand, several investigations have demonstrated increased 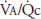 inequality for the whole lung during exercise. The mechanism for this is unclear, but it may involve increased perfusion of poorly ventilated alveoli. This may occur as increased pulmonary artery pressure interferes with hypoxic pulmonary vasoconstriction (HPV). Increased perfusion of poorly ventilated alveoli would act as an intrapulmonary shunt (or a “shunt-like” area). This is one explanation for the decrease in arterial
inequality for the whole lung during exercise. The mechanism for this is unclear, but it may involve increased perfusion of poorly ventilated alveoli. This may occur as increased pulmonary artery pressure interferes with hypoxic pulmonary vasoconstriction (HPV). Increased perfusion of poorly ventilated alveoli would act as an intrapulmonary shunt (or a “shunt-like” area). This is one explanation for the decrease in arterial 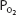 and increased alveolar-arterial
and increased alveolar-arterial 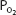 difference (the (A-a)
difference (the (A-a)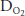 ) seen in normal people as exercise levels increase. The (A-a)
) seen in normal people as exercise levels increase. The (A-a)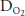 , which is normally 5 to 15 mm Hg at rest in a young person, usually increases to 20 to 30 mm Hg in normal healthy untrained people at maximal exercise and may be even greater in athletes. Another explanation for the decreased arterial
, which is normally 5 to 15 mm Hg at rest in a young person, usually increases to 20 to 30 mm Hg in normal healthy untrained people at maximal exercise and may be even greater in athletes. Another explanation for the decreased arterial 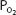 and the increased (A-a)
and the increased (A-a)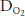 during strenuous exercise is that intrapulmonary arteriovenous shunts may open as exercise levels increase. These intrapulmonary arteriovenous shunts, which appear to anatomically bypass pulmonary capillaries, may open to help keep pulmonary vascular pressures low to prevent pulmonary edema caused by high pulmonary capillary hydrostatic pressure or vessel failure caused by mechanical stress (“stress failure”). Lack or dysfunction of these shunts may be involved in the development of high-altitude pulmonary edema (see the next section of this chapter) in some people, while others at the same altitude don’t have problems with it. Furthermore, as exercise levels increase, mixed venous
during strenuous exercise is that intrapulmonary arteriovenous shunts may open as exercise levels increase. These intrapulmonary arteriovenous shunts, which appear to anatomically bypass pulmonary capillaries, may open to help keep pulmonary vascular pressures low to prevent pulmonary edema caused by high pulmonary capillary hydrostatic pressure or vessel failure caused by mechanical stress (“stress failure”). Lack or dysfunction of these shunts may be involved in the development of high-altitude pulmonary edema (see the next section of this chapter) in some people, while others at the same altitude don’t have problems with it. Furthermore, as exercise levels increase, mixed venous 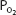 decreases (see below), which would also increase the effect on the (A-a)
decreases (see below), which would also increase the effect on the (A-a) . Finally, the development of diffusion limitation of oxygen transfer (see the next section) from the alveolus to the pulmonary capillary may contribute to the increased (A-a)
. Finally, the development of diffusion limitation of oxygen transfer (see the next section) from the alveolus to the pulmonary capillary may contribute to the increased (A-a) in strenuous exercise.
in strenuous exercise.
Figure 11–1.
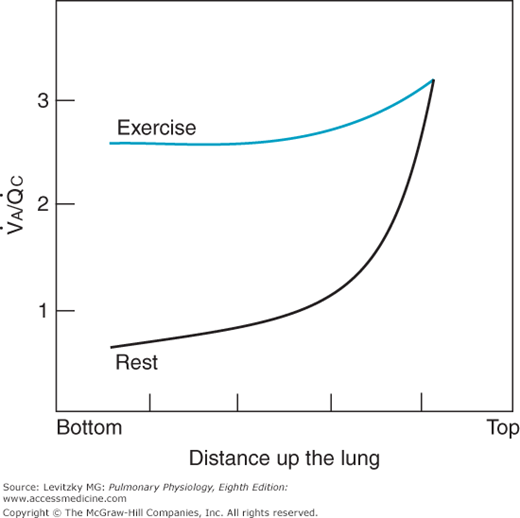
Representation of regional differences in the ventilation-perfusion ratio of an upright lung during rest and exercise. Compare with Figure 5–7.
The diffusing capacities for oxygen and carbon dioxide normally increase substantially during exercise. Some studies have shown a nearly linear increase in diffusing capacity as oxygen uptake increases, although the diffusing capacity may reach a maximum level before the 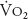 does. The increase in diffusing capacity during exercise is largely a result of the increase in pulmonary blood flow. Recruitment of capillaries, especially in upper regions of the lungs, increases the surface area available for diffusion. Increased linear velocity of blood flow through pulmonary capillaries reduces the time that red blood cells spend in contact with the alveolar air to less than the 0.75 seconds normally seen at rest, decreasing the perfusion limitation of gas transfer. As noted in Chapter 6, the
does. The increase in diffusing capacity during exercise is largely a result of the increase in pulmonary blood flow. Recruitment of capillaries, especially in upper regions of the lungs, increases the surface area available for diffusion. Increased linear velocity of blood flow through pulmonary capillaries reduces the time that red blood cells spend in contact with the alveolar air to less than the 0.75 seconds normally seen at rest, decreasing the perfusion limitation of gas transfer. As noted in Chapter 6, the 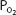 and
and 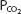 of the plasma in the pulmonary capillaries normally equilibrate with the alveolar
of the plasma in the pulmonary capillaries normally equilibrate with the alveolar 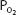 and
and 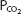 within about the first 0.25 seconds of the time blood spends in the pulmonary capillaries. After this equilibration, no further gas diffusion between the equilibrated blood and the alveoli takes place because the partial pressure gradient (P1 – P2 in Fick law) is equal to zero. Increased velocity of blood flow through the lung therefore increases the diffusing capacity by bringing unequilibrated blood into the lung faster, thus maintaining the partial pressure gradient for diffusion. On the other hand, very great blood flow velocities may increase the possibility of diffusion limitation of gas transfer, even in a healthy person. Diffusion limitation is likely to develop in a person who has some diffusion impairment, as can be seen in Figure 6–2.
within about the first 0.25 seconds of the time blood spends in the pulmonary capillaries. After this equilibration, no further gas diffusion between the equilibrated blood and the alveoli takes place because the partial pressure gradient (P1 – P2 in Fick law) is equal to zero. Increased velocity of blood flow through the lung therefore increases the diffusing capacity by bringing unequilibrated blood into the lung faster, thus maintaining the partial pressure gradient for diffusion. On the other hand, very great blood flow velocities may increase the possibility of diffusion limitation of gas transfer, even in a healthy person. Diffusion limitation is likely to develop in a person who has some diffusion impairment, as can be seen in Figure 6–2.
Another related factor, less dependent on the increased cardiac output, that helps increase diffusion during exercise is that the mixed venous 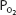 may be lower and the mixed venous
may be lower and the mixed venous 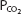 may be higher than those seen at rest. These factors may also help increase and maintain the partial pressure gradients for diffusion.
may be higher than those seen at rest. These factors may also help increase and maintain the partial pressure gradients for diffusion.
The total effect on diffusion through the alveolar-capillary barrier of the increased surface area and the better maintenance of the alveolar-capillary partial pressure gradients may be seen by reviewing Fick law for diffusion:
The thickness of the alveolar-capillary barrier may also be affected during exercise, but the net effect may be either an increase or a decrease. At high lung volumes, the alveolar vessels are stretched and the thickness of the barrier may decrease. On the other hand, high cardiac outputs may be associated with vascular congestion, increasing the thickness of the barrier.
The alveolar-arterial oxygen difference increases during exercise. This is probably because of a number of factors, including 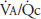 mismatch, diffusion limitation of gas transfer, a decreased mixed venous
mismatch, diffusion limitation of gas transfer, a decreased mixed venous 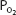 , an increased alveolar
, an increased alveolar 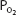 , and alterations in the oxyhemoglobin dissociation curve.
, and alterations in the oxyhemoglobin dissociation curve.
The loading of carbon dioxide into the blood and the unloading of oxygen from the blood are enhanced in exercising muscles. Oxygen unloading is improved because the 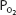 in the exercising muscle is decreased, causing a larger percentage of deoxyhemoglobin. Oxygen unloading is also enhanced by the rightward shift of the oxyhemoglobin dissociation curve caused by the elevated
in the exercising muscle is decreased, causing a larger percentage of deoxyhemoglobin. Oxygen unloading is also enhanced by the rightward shift of the oxyhemoglobin dissociation curve caused by the elevated 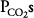 (the Bohr effect), hydrogen ion concentrations, and temperatures (and possibly 2,3-bisphosphoglycerate [2,3-BPG]) found in exercising muscle. Low capillary
(the Bohr effect), hydrogen ion concentrations, and temperatures (and possibly 2,3-bisphosphoglycerate [2,3-BPG]) found in exercising muscle. Low capillary 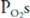 should also lead to improved carbon dioxide loading because lower oxyhemoglobin levels shift the carbon dioxide dissociation curve to the left (the Haldane effect).
should also lead to improved carbon dioxide loading because lower oxyhemoglobin levels shift the carbon dioxide dissociation curve to the left (the Haldane effect).
Exercise strenuous enough to cause a significant degree of anaerobic metabolism results in metabolic acidosis secondary to the increased lactic acid production. As discussed previously, the hydrogen ions generated by this process stimulate the arterial chemoreceptors (especially the carotid bodies) and cause a further compensatory increase in alveolar ventilation, maintaining arterial pH near the normal level.
The responses of the normal respiratory system to acute exercise are summarized in Table 11–1.
| Variable | Moderate Exercise | Strenuous Exercise |
|---|---|---|
| Mechanics of breathing | ||
| Elastic work of breathing | ↑ | ↑ ↑ |
| Resistance work of breathing | ↑ | ↑ ↑ |
| Alveolar ventilation | ||
| Tidal volume | ↑ ↑ | ↑ ↑ |
| Frequency | ↑ | ↑ ↑ |
| Anatomic dead space | ↑ | ↑ |
| Alveolar dead space (if present) | ↓ | ↓ |
| Vd/Vt | ↓ | ↓ ↓ |
| Pulmonary blood flow | ↑ | ↑ ↑ |
| Perfusion of upper lung | ↑ | ↑ ↑ |
| Pulmonary vascular resistance | ↓ | ↓ ↓ |
| Linear velocity of blood flow | ↑ | ↑ ↑ |
| Ventilation-perfusion relationships | ||
| Ventilation-perfusion matching | ↑ | ↑ |
| Ventilation-perfusion ratio | ↑ | ↑ ↑ |
| Diffusion through the alveolar-capillary barrier | ||
| Surface area | ↑ | ↑ ↑ |
| Perfusion limitation | ↓ | ↓ ↓ |
| Partial pressure gradients | ↑ | ↑ ↑ |
| Oxygen unloading at the tissues | ↑ | ↑ ↑ |
| Carbon dioxide loading at the tissues | ↑ | ↑ ↑ |
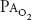 | ↔ | ↑ |
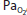 | ↔ | ↑, ↔ or ↓ |
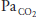 | ↔ | ↓ |
| pHa | ↔ | ↓ |
| Arteriovenous O2 difference | ↑ | ↑ ↑ |
The ability to perform physical exercise increases with training. Most of the changes that occur as a result of physical training, however, are a function of alterations in the cardiovascular system and in muscle metabolism rather than changes in the respiratory system. The maximal oxygen uptake increases with physical training. This increase appears to be mainly a result of an increased maximal cardiac output. As stated earlier in this chapter, the maximal cardiac output is probably a limiting factor in exercise. Physical training lowers the resting heart rate and increases the resting stroke volume. The maximal heart rate does not appear to be affected by physical training, but the heart rate of a trained person is lower than that of an untrained person at any level of physical activity. Stroke volume is increased. The arterial hemoglobin concentration and the hematocrit do not appear to change with physical training at sea level, but the arteriovenous oxygen content difference does appear to increase with physical training. This is probably a function of the increased effects of local pH, 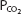 , and temperature in the exercising muscles, as well as an increased ability of the muscles to use oxygen. Blood volume is usually increased by training.
, and temperature in the exercising muscles, as well as an increased ability of the muscles to use oxygen. Blood volume is usually increased by training.
Physical training increases the oxidative capacity of skeletal muscle by inducing mitochondrial proliferation and increasing the concentration of oxidative enzymes and the synthesis of glycogen and triglyceride. These alterations result in lower concentrations of blood lactate in trained subjects than those found in untrained people, reflecting increased aerobic energy production. Nevertheless, blood lactate levels during maximal exercise may be greater in trained athletes than in untrained people.
Maximal ventilation and resting ventilation do not appear to be affected by physical training, but ventilation at submaximal loads is decreased, probably because of the lower lactic acid levels of the trained person during submaximal exercise. The strength and endurance of the respiratory muscles appear to improve with training. Total lung capacity is not affected by training; vital capacity may be normal or elevated. Pulmonary diffusing capacity is often elevated in athletes, probably as a result of their increased blood volumes and maximal cardiac outputs.
Altitude and Acclimatization
The total barometric pressure decreases at greater altitudes because the total barometric pressure at any altitude is proportional to the weight of the air above it. There is a greater change in barometric pressure per change in altitude closer to the earth’s surface than there is at very great altitudes because air, which is attracted to the earth’s surface by gravity, is compressible.
The fractional concentration of oxygen in the atmosphere does not change appreciably with altitude. Oxygen constitutes about 21% of dry ambient air, and so the 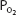 of dry air at any altitude is about 0.21 times total barometric pressure at that altitude. Water vapor pressure must also be considered in calculations of the
of dry air at any altitude is about 0.21 times total barometric pressure at that altitude. Water vapor pressure must also be considered in calculations of the 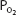 . The water vapor pressure depends on the temperature and humidity of the air. As the inspired air passes through the airways, it is normally warmed to body temperature and completely humidified. Therefore, the partial pressure exerted by the water vapor in the air entering the alveoli is fixed at 47 mm Hg.
. The water vapor pressure depends on the temperature and humidity of the air. As the inspired air passes through the airways, it is normally warmed to body temperature and completely humidified. Therefore, the partial pressure exerted by the water vapor in the air entering the alveoli is fixed at 47 mm Hg.
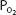
The inspired 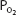 is equal to 0.21 times the total barometric pressure (if ambient air is breathed) after the subtraction of the water vapor pressure of 47 mm Hg:
is equal to 0.21 times the total barometric pressure (if ambient air is breathed) after the subtraction of the water vapor pressure of 47 mm Hg:
The alveolar 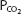 falls at greater altitudes because hypoxic stimulation of the arterial chemoreceptors increases alveolar ventilation. For example, at an altitude of 15,000 ft (4572 m), the total barometric pressure is about 429 mm Hg. The inspired
falls at greater altitudes because hypoxic stimulation of the arterial chemoreceptors increases alveolar ventilation. For example, at an altitude of 15,000 ft (4572 m), the total barometric pressure is about 429 mm Hg. The inspired 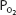 is therefore 0.21 × (429 − 47) mm Hg, or 80.2 mm Hg. The alveolar
is therefore 0.21 × (429 − 47) mm Hg, or 80.2 mm Hg. The alveolar 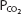 is likely to be decreased to about 32 mm Hg, resulting in a
is likely to be decreased to about 32 mm Hg, resulting in a 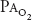 of about 45 mm Hg. At 18,000 ft (5486 m), the total barometric pressure is about 380 mm Hg; at 20,000 ft (6096 m), it is 349 mm Hg. At 50,000 ft (15,240 m), the total barometric pressure is only 87 mm Hg. Even if 100% oxygen is breathed, the
of about 45 mm Hg. At 18,000 ft (5486 m), the total barometric pressure is about 380 mm Hg; at 20,000 ft (6096 m), it is 349 mm Hg. At 50,000 ft (15,240 m), the total barometric pressure is only 87 mm Hg. Even if 100% oxygen is breathed, the 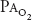 plus the
plus the 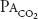 divided by R can only total 40 mm Hg after water vapor pressure is subtracted. At 63,000 ft, the total barometric pressure is 47 mm Hg and the fluid in blood “boils.”
divided by R can only total 40 mm Hg after water vapor pressure is subtracted. At 63,000 ft, the total barometric pressure is 47 mm Hg and the fluid in blood “boils.”
The advantage of hypoxic stimulation of the arterial chemoreceptors causing the hyperventilation described in the previous paragraph can be understood by looking at Figure 11–2 and considering the alveolar air equation. As the alveolar 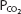 decreases with increasing alveolar ventilation, the alveolar
decreases with increasing alveolar ventilation, the alveolar 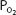 is closer to the inspired
is closer to the inspired 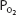 at that altitude. The cost is respiratory alkalosis (as well as increased work of breathing, of course).
at that altitude. The cost is respiratory alkalosis (as well as increased work of breathing, of course).
Figure 11–2.
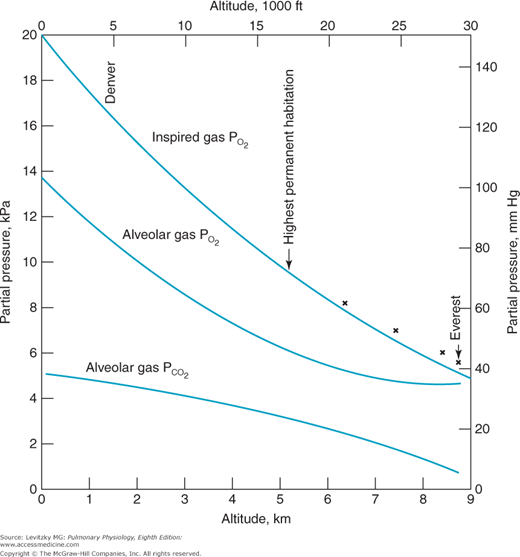
Calculated inspired and alveolar partial pressures of oxygen and carbon dioxide at rest plotted versus increasing altitude. Note that as increasing arterial chemoreceptor drive decreases alveolar 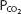 , alveolar
, alveolar 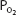 is closer to inspired
is closer to inspired 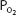 . (Adapted with permission from Lumb AB. Nunn’s Applied Respiratory Physiology. 5th ed. Oxford: Butterworth-Heinemann; 2000 [Figure 16.1, p. 359].)
. (Adapted with permission from Lumb AB. Nunn’s Applied Respiratory Physiology. 5th ed. Oxford: Butterworth-Heinemann; 2000 [Figure 16.1, p. 359].)
An unacclimatized person would suffer a deterioration of nervous system function if he or she ascended rapidly to great heights. Similar dysfunctions occur if cabin pressure is lost in an airplane. The symptoms are mainly due to hypoxia and may include sleepiness, laziness, a false sense of well-being, impaired judgment, blunted pain perception, increasing errors on simple tasks, decreased visual acuity, clumsiness, and tremors. Severe hypoxia, of course, may result in a loss of consciousness or even death.
If an unacclimatized person ascends to a moderate altitude (8000–10,000 ft or 2400–3000 m above sea level), he or she may suffer from a group of symptoms known collectively as acute mountain sickness.  The symptoms include headache, dizziness, breathlessness at rest, weakness, malaise, nausea, anorexia, sweating, palpitations, dimness of vision, partial deafness, sleeplessness, fluid retention, and dyspnea on exertion. These symptoms are a result of hypoxia and hypocapnia, and alkalosis or cerebral edema, or both.
The symptoms include headache, dizziness, breathlessness at rest, weakness, malaise, nausea, anorexia, sweating, palpitations, dimness of vision, partial deafness, sleeplessness, fluid retention, and dyspnea on exertion. These symptoms are a result of hypoxia and hypocapnia, and alkalosis or cerebral edema, or both.
The decreased alveolar and arterial 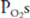 that occur at altitude result in stimulation of the arterial chemoreceptors and an increase in alveolar ventilation; the central chemoreceptors are not responsive to hypoxia. At an arterial
that occur at altitude result in stimulation of the arterial chemoreceptors and an increase in alveolar ventilation; the central chemoreceptors are not responsive to hypoxia. At an arterial 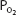 of 45 mm Hg, minute ventilation is approximately doubled. Because carbon dioxide production is initially normal (it does increase with the elevated work of breathing caused by greater alveolar ventilation), alveolar and arterial
of 45 mm Hg, minute ventilation is approximately doubled. Because carbon dioxide production is initially normal (it does increase with the elevated work of breathing caused by greater alveolar ventilation), alveolar and arterial 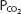 fall, causing respiratory alkalosis. Arterial hypocapnia also results in “diffusion” of carbon dioxide out of the cerebrospinal fluid, causing an increase in the pH of the cerebrospinal fluid. The central chemoreceptors are therefore not only unresponsive to the hypoxia of altitude; their activity is depressed by the secondary hypocapnia and alkalosis of the cerebrospinal fluid.
fall, causing respiratory alkalosis. Arterial hypocapnia also results in “diffusion” of carbon dioxide out of the cerebrospinal fluid, causing an increase in the pH of the cerebrospinal fluid. The central chemoreceptors are therefore not only unresponsive to the hypoxia of altitude; their activity is depressed by the secondary hypocapnia and alkalosis of the cerebrospinal fluid.
The increased rate and depth of breathing increase the work of breathing. Greater transpulmonary pressures are necessary to generate greater tidal volumes and also to overcome the possible effects of vascular engorgement and increased interstitial fluid volume of the lung, which may also decrease the vital capacity during the first 24 hours at altitude. High ventilatory rates may be accompanied by active expiration, resulting in dynamic compression of airways. This airway compression, coupled with a reflex parasympathetic bronchoconstriction in response to the arterial hypoxemia, results in elevated resistance work of breathing. More turbulent airflow, which is likely to be encountered at elevated ventilatory rates, may also contribute to elevated resistance work. Maximum airflow rates may increase because of decreased gas density.
The anatomic dead space may decrease slightly at altitude because of the reflex bronchoconstriction or increase slightly because of the opposing effect of increased tidal volumes. In any event the ratio of dead space to tidal volume falls with greater tidal volumes. A more uniform regional distribution of alveolar ventilation is also expected at altitude because of deeper inspirations and fuller expirations. Previously collapsed or poorly ventilated alveoli will be better ventilated.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


