Chapter Twenty-Seven. The peripheral and autonomic nervous systems
CHAPTER CONTENTS
The peripheral nervous system
Introduction
The peripheral nervous system (PNS) detects changes in the body’s external or internal environments. Sensory receptors code them into nerve impulses and pass the information back to the central nervous system (CNS) so that appropriate action can occur. Some messages are not passed to the brain; they influence reflex actions at the level of the spinal cord or brainstem. The PNS includes all the neural structures outside the brain and spinal cord (i.e. sensory receptors), peripheral nerves and associated ganglia and efferent motor endings (Marieb 2008). For ease of understanding of nerve pathways to the brain, the spinal cord columns are placed in this chapter.
Ascending sensory tracts
Categories of sensation
There are two kinds of sensation (Fig. 27.1): conscious sensations perceived at the level of the cortex and non-conscious sensations that are not. Conscious sensation can be divided into exteroception and proprioception. Exteroception involves messages from the outside world perceived in the cerebral cortex. Sensations may originate in body surface receptors or in telereceptors of the special senses such as vision or hearing. Proprioceptors in the locomotor system and the inner ear labyrinth inform the brain of the position when stationary ( position sense) and during movement ( kinaesthetic sense).
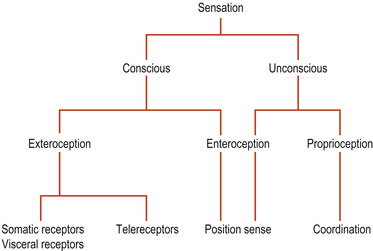 |
| Figure 27.1 Categories of sensation. |
Non-conscious sensation can also be divided into two kinds: non-conscious proprioception, which affects the cerebellum, involving messages essential for smooth muscle coordination received through spinocerebellar pathways and the brainstem, and enteroception which refers to non-conscious signals from visceral reflexes (Fitzgerald et al 2006).
Somatic sensory perception
Two major pathways are involved in somatic perception sensations: the posterior column–medial lemniscal pathway and the spinothalamic pathway (Fig. 27.2). There are common features:
• They contain first-order, second-order and third-order sensory neurons.
• The cell bodies of the first-order neurons are in the posterior root ganglia.
• The cell bodies of the second-order neurons are on the same side of the CNS grey matter as the first-order neurons.
• Second-order axons cross the midline to ascend and terminate in the thalamus.
• Third-order neurons project to the somatosensory cortex.
• Both pathways are somatotopic, representing the body parts in an orderly fashion up to the sensory cortex.
• Both pathways can be modulated, either by inhibition or stimulation by other neurons.
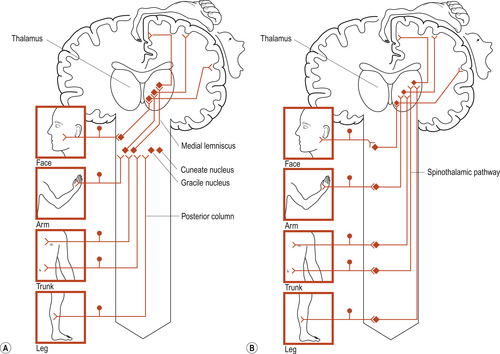 |
| Figure 27.2 Basic plans of (A) posterior column–medial lemniscal pathway, (B) spinothalamic pathway. (From Fitzgerald M J T 1996, with permission.) |
The posterior column–medial lemniscal pathway
The first-order nerve fibres enter the dorsal columns of the spinal cord without synapsing. They are usually large A fibres with conduction velocities of about 70 metres per second (m/s). As nerve fibres from higher levels in the cord are added, they take up lateral positions so that the higher the level of origin is, the more lateral the position of the fibre in the column. The fibres of second-order neurons at the level of the brainstem cross the midline to be projected to the thalamus. Crossing over is why one side of the brain controls the opposite (contralateral) side of the body. Cells in the sensory relay nucleus of the thalamus are third-order neurons and project their fibres to the somatosensory cortex.
The chief functions of this pathway are conscious proprioception and discriminatory touch. These provide the parietal lobe with an instantaneous body image of our position both at rest and when moving. Disturbances of this pathway cause demyelinating diseases such as multiple sclerosis.
The spinothalamic tract
The dorsal root fibres of this pathway tend to be the smaller A-δ or unmyelinated C fibres with slow conduction velocity. The dorsal root fibres enter the spinal cord and may ascend or descend a few segments before synapsing with cells of the dorsal horn in the substantia gelatinosa. The dorsal horn second-order fibres ascend or descend a few segments before crossing over the midline to ascend in the spinothalamic tract. These fibres terminate on thalamic third-order neurons whose fibres synapse on cells of the sensory cortex. The role of this pathway is the perception of heat, cold and touch on the opposite side of the body. The role of the substantia gelatinosa in the gate control theory of pain perception is discussed in Chapter 38.
Somatosensory receptors
Sensory receptors are mostly adapted nerve fibre endings that respond to environmental changes. Sensory afferent nerves arising from the body are grouped together as the somatosensory system and include sensation from the skin, muscles, joints and viscera. The special senses are associated with organs in the head and include vision, hearing, balance, taste and smell; they are not discussed in this book but a useful source is Marieb (2008).
Types of somatosensory receptors
• Mechanoreceptors respond to touch, pressure, vibrations and stretch.
• Thermoreceptors respond to temperature change.
• Photoreceptors respond to light.
• Chemoreceptors respond to smell, taste and changes in blood chemistry.
• Nociceptors respond to damage by causing pain (see Ch. 38).
Descending motor pathways
The descending tracts carry efferent messages from the brain down the spinal cord and are divided into four main pathways:
1. Corticospinal (pyramidal).
2. Reticulospinal (extrapyramidal).
3. Vestibulospinal.
4. Tectospinal.
The corticospinal tract
The corticospinal tract (Fig. 27.3) is the major motor pathway involved with voluntary movement. It contains about 1 million nerve fibres, more than 60% of which originate in the primary motor cortex. The tract descends through the internal capsule to the brainstem. It continues through the pons to the medulla oblongata where about 80% of the fibres decussate (cross over to the other side of the body). The fibres, which are arranged somatotopically (Fig. 27.4), synapse with interneurons or directly with anterior horn neurons.
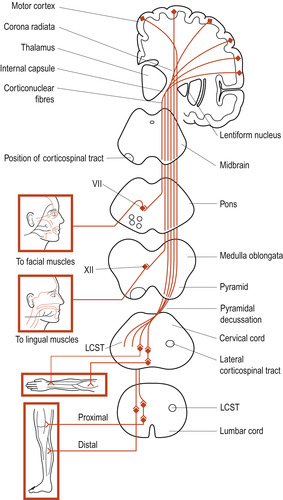 |
| Figure 27.3 Corticospinal tract viewed from the front. At spinal cord level, only the lateral corticospinal tract is shown. LCST, lateral corticospinal tract; VII, nucleus of facial nerve; XII, hypoglossal nucleus. (From Fitzgerald M J T 1996, with permission.) |
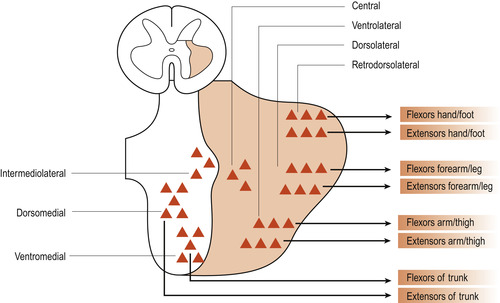 |
| Figure 27.4 Cell columns in the anterior grey horn of the spinal cord: somatotopic organisation. (From Fitzgerald M J T 1996, with permission.) |
The reticulospinal tract
The reticulospinal tract is partially crossed and originates in the reticular formation of the pons and medulla. It is involved in two kinds of motor behaviour: locomotion, where it controls bilateral rhythmicity (try moving the arm and leg of the same side together when walking), and postural control.
The vestibulospinal tract
The vestibulospinal tract is an uncrossed paired pathway originating in the vestibular nucleus of the medulla oblongata. It maintains balance when the head is tilted to one side.
The tectospinal tract
The tectospinal tract is a crossed pathway descending from the tectum of the midbrain to the medial part of the anterior horn at cervical and upper thoracic levels. In reptiles it orients the head and trunk towards visual or auditory stimuli and may have a similar function in humans.
Upper and lower motor neurons
The neurons of the motor cortex are called the pyramidal cells because of the shape of their bodies. They are referred to as the upper motor neurons. The anterior horn neurons whose axons leave the cord to innervate skeletal muscles are called the lower motor neurons. Damage to upper motor neurons causes floppy paralysis with loss of tendon reflexes. Damage to lower motor neurons causes weakness and wasting of muscles. Motor neuron disease is characterised by progressive degeneration of both upper and lower motor neurons (Fitzgerald et al 2006).



