Chapter Five. The male reproductive system
Anatomy of the male reproductive system
The male genitalia are mainly outside the body cavity, a situation necessary for both production and transfer of spermatozoa. The organs are the scrotum, testis, rete and epididymis, ductus deferens, seminal vesicles, prostate gland, bulbourethral glands and penis with the urethra (Fig. 5.1). Unlike the female urinary system where the urethral orifice is separate to the vagina, the male genital and urinary systems share a common outlet through the urethra.
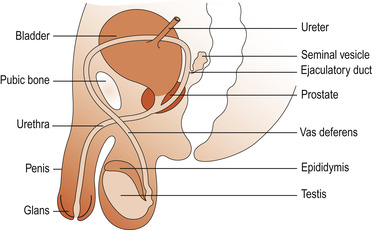 |
| Figure 5.1 The male reproductive system. (From Henderson C, Macdonald S 2004, with kind permission of Elsevier.) |
The scrotum and testes
Embryonic development
As discussed in Chapter 4, SRY activity on the Y chromosome converts the indifferent gonad to a testis. In the absence of this factor the gonad develops into an ovary (Jones 2002). It is an efficient process and there are few true hermaphrodites who have both testicular and ovarian tissue. Once the gonad is established the SRY gene is switched off. The testicular Sertoli cells (see below) also secrete Müllerian inhibiting hormone (MIH), which remains active until puberty, when there is a rapid decline in function. The Leydig cells (see below) of the testis produce testosterone from 13–15 weeks (Johnson 2007).
In the embryo, the testes develop high up on the lumbar region of the abdominal cavity. In the last few months of fetal life they descend through the abdominal cavity, over the pelvic brim and down the inguinal canal into the scrotal sac outside the body cavity. This descent occurs under the influence of testosterone and is completed in 98% of boys by birth.
The mature testis
At maturity each testis measures 4 cm long and 3 cm in diameter and is surrounded by two coats: the outer tunica vaginalis, which is derived from peritoneum, and the inner fibrous capsule, the tunica albuginea. One testis sits in each pocket of the scrotal sac. The scrotum is a thin-walled sac covered with hairy, rugose skin well supplied with sebaceous glands. The scrotal skin is highly vascularised and has a large surface area.
The temperature of the testes is maintained at 2–3°C below that of the body core to facilitate spermatogenesis. The position of the scrotum relative to the body can be adjusted by a spinal reflex in order to regulate testicular temperature. In a cold environment contraction of the scrotal muscle, the dartos muscle, wrinkles the scrotal skin and reduces the size of the sac, whereas the cremaster muscle, a skeletal muscle arising from the internal oblique muscle, contracts and lifts the testes nearer to the body. Relaxation of these muscles allows the testes to be held away from the body to facilitate cooling.
Structure
Each testis is divided into 200–300 wedge-shaped lobules by thin fibrous partitions that are extensions of the tunica albuginea (Fig. 5.2). Each lobule contains up to four seminiferous tubules which are highly coiled loops. About 80% of the testis by weight consists of seminiferous tubules in which spermatozoa develop. The seminiferous tubules of each lobule converge to form a straight tubule or tubulus rectus that conveys the sperm into the rete testis, a tubular network on the posterior aspect of the testis. From here the sperm enter the epididymis which is in close apposition to the external surface of the testis. Macrophages that phagocytose dead sperm are found in the lumen of the epididymis. Interstitial tissue is packed around the seminiferous tubules and contains blood vessels and endocrine cells, the Leydig cells, which secrete testosterone.
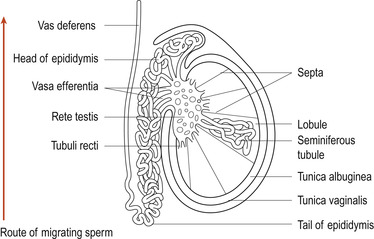 |
| Figure 5.2 The testis. (From Hinchliff S M, Montague S E 1990, with kind permission of Elsevier.) |
Blood supply
The testes are supplied by the testicular arteries that arise from the abdominal aorta. The testicular veins form a network around the testicular artery called a pampiniform plexus (tendril-like). This absorbs heat from the artery before the blood enters the testis.
Lymphatic drainage
Lymphatic drainage is by the inguinal nodes.
Nerve supply
There is innervation by the autonomic system—both sympathetic and parasympathetic. There is also a rich sensory nerve supply, resulting in much pain and nausea if the testes are struck. The nerve fibres run with the blood vessels and lymphatics in the fibrous connective tissue sheath called the spermatic cord.
Function of the testes
• To produce spermatozoa.
• To produce the hormones testosterone and inhibin.
Spermatogenesis (the production of spermatozoa)
Each testis consists of two separate compartments: cells that produce sperm and those that produce hormones. There is a physical barrier consisting of cellular barriers which limit free exchange of water-soluble materials. This barrier develops at puberty and is formed of multiple layers of gap and tight junctions surrounding each Sertoli cell. This is the blood–testis barrier (Johnson 2007). Its functions are:
1. To prevent sperm from entering the systemic and lymphatic circulations where they could set up antisperm antibodies, leading to infertility.
2. To maintain distinct chemical environments on either side of the barrier to facilitate sperm development and health.
Seminiferous tubules contain two types of cell: germ cells and Sertoli cells (Fig. 5.3). Primary germ cells are dormant from the fetal period of life and begin to increase in number at puberty. Sperm production in the seminiferous tubules has three phases:
1. Mitotic proliferation, which produces large numbers of cells.
2. Meiotic division, which generates diversity and halves the chromosome number (haploid).
3. Cytodifferentiation, which packages the chromosomes for effective delivery.
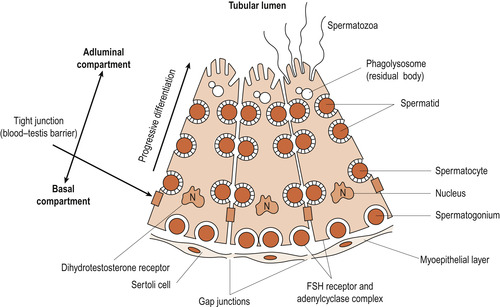 |
| Figure 5.3 Sertoli cells with developing germ cells at all stages of development from spermatogonia to spermatozoa. (From Tepperman & Tepperman 1987, with permission.) |
In a functioning testis, germ cells will be present at all stages of development, all originating from spermatogonia. Spermatogonia divide by mitosis continuously to ensure a constant supply of cells maturing towards sperm. After undergoing several mitotic divisions they mature, become larger and are known as primary spermatocytes but are still diploid cells. Although nuclear division ( karyokinesis) occurs, cytoplasmic division is incomplete and spermatogonia are linked by cytoplasmic bridges to form a syncytium. This persists throughout the meiotic phase and individual cells are only released as mature sperm (Johnson 2007).
Primary spermatocytes undergo the first meiotic division to form two secondary spermatocytes which have only 23 chromosomes—one of each pair. Half will receive the X chromosome and half the Y chromosome. Secondary meiosis results in four haploid cells called spermatids.
Spermatids are found in close association with Sertoli cells, which are polymorphic cells attached to a basement membrane but extending into the lumen of the seminiferous tubule. They provide nutrition and support to the sperm and are sometimes called ‘nurse cells’. Here the spermatids are transformed from fairly basic cells into highly specialised sperm.
As a sperm matures, excess protoplasm is lost and the chromatin of the nucleus condenses to become the head. One centriole develops into the tail, which is composed of a central filament of two microfibrils surrounded by a circle of nine fibrils. Mitochondria aggregate into the neck region and the Golgi apparatus helps to form the acrosome cap which develops over the head of the sperm and contains enzymes called hyaluronidases and proteases.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


