CHAPTER 27 The Cephalic, Oral, and Esophageal Phases of the Integrated Response to a Meal
In this chapter we will look at the processes that occur in the gastrointestinal (GI) tract in the early stages of the integrated response to a meal. Even before food is ingested there are changes in the physiology of the GI tract, and in this so-called cephalic phase, as well as in the oral phase (when ingested food is in the mouth), the responses of the GI tract to the presence of food are mainly associated with preparing the GI tract for digestion and absorption. We will also look at the transfer of food from the mouth to the esophagus, the esophageal phase of the meal.
CEPHALIC AND ORAL PHASES
Properties of Secretion
General Considerations
Secretions in the GI tract come from glands associated with the tract (the salivary glands, pancreas, and liver), from glands formed by the gut wall itself (e.g., Brunner’s glands in the duodenum), and from the intestinal mucosa itself. The exact nature of the secretory products can vary tremendously, depending on the function of that region of the GI tract. However, these secretions have several characteristics in common. Secretions from the GI tract and associated glands include water, electrolytes, protein, and humoral agents. Water is essential for generating an aqueous environment for the efficient action of enzymes. Secretion of electrolytes is important for the generation of osmotic gradients to drive the movement of water. Digestive enzymes in secreted fluid catalyze the breakdown of macronutrients in ingested food. Moreover, many additional proteins secreted along the GI tract have specialized functions, some of which are fairly well understood, such as those of mucin and immunoglobulins, and others that are only just beginning to be understood, such as those of trefoil peptides.
Salivary Secretion
During the cephalic and the oral phase of the meal, considerable stimulation of salivary secretion takes place. Saliva has a variety of functions, including those important for the integrative responses to a meal and for other physiological processes (Table 27-1). The main functions of saliva in digestion include lubrication and moistening of food for swallowing, solubilization of material for taste, initiation of carbohydrate digestion, and clearance and neutralization of refluxed gastric secretions in the esophagus. Saliva also has antibacterial actions that are important for overall health of the oral cavity and teeth.
Table 27-1 Functions of Saliva and Chewing
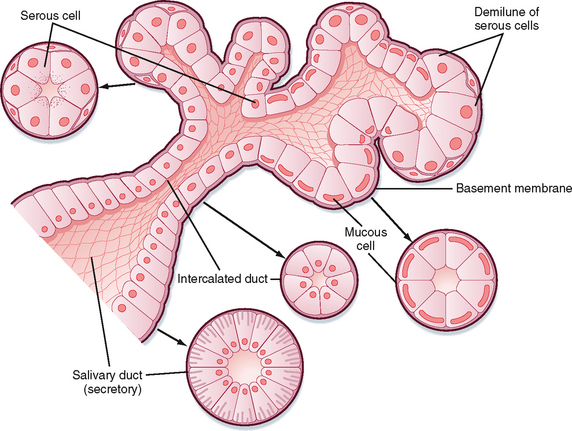
Functional Anatomy of the Salivary Glands
There are three pairs of major salivary glands: parotid, submandibular, and sublingual. In addition, many smaller glands are found on the tongue, lips, and palate. These glands are the typical tubuloalveolar structures of glands located in the GI tract (Fig. 27-1). The acinar portion of the gland is classified according to its major secretion: serous (“watery”), mucous, or mixed. The parotid gland produces mainly serous secretion, the sublingual gland secretes mainly mucus, and the submandibular gland produces a mixed secretion.
Composition of Saliva
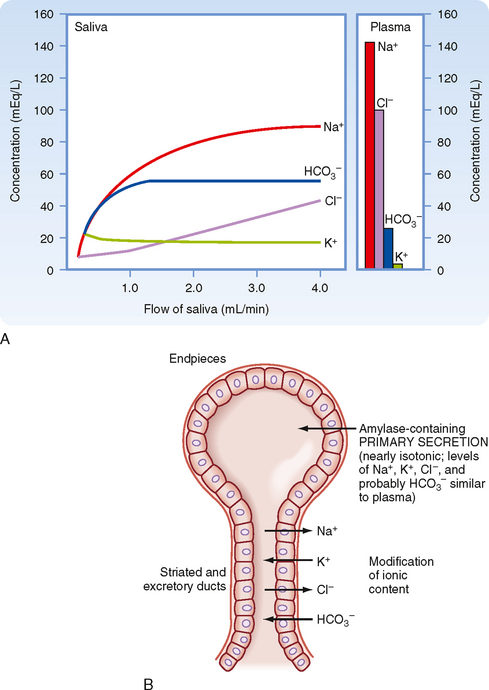
The primary secretion is produced by acinar cells in the secretory end pieces (acini) and is modified by duct cells as saliva passes through the ducts. The primary secretion is isotonic, and the concentration of the major ions is similar to that in plasma. Secretion is driven predominantly by Ca++-dependent signaling, which opens apical Cl− channels in the acinar cells. Cl− therefore flows out into the duct lumen and establishes an osmotic and electrical gradient. Because the epithelium of the acinus is relatively leaky, Na+ and water then follow across the epithelium via the tight junctions (i.e., via paracellular transport). Transcellular water movement may also occur, mediated by aquaporin 5 water channels. The amylase content and rate of fluid secretion vary with the type and level of stimulus. The excretory duct cells and the striated duct cells modify the primary secretion to produce the secondary secretion. The duct cells reabsorb Na+ and Cl− and secrete K+ and HCO3− into the lumen. At rest, the final salivary secretion is hypotonic and slightly alkaline. Na+ is exchanged for protons, but some of the secreted protons are then reabsorbed in exchange for K+. HCO3−, on the other hand, is secreted only in exchange for Cl−, thereby providing excess base equivalents. The alkalinity of saliva is probably important in restricting microbial growth in the mouth, as well as in neutralizing refluxed gastric acid once the saliva is swallowed. When salivary secretion is stimulated, moreover, there is a decrease in K+ (but it always remains above plasma concentrations), Na+ increases toward plasma levels, Cl− and HCO3− increase, and thus the secretion becomes even more alkaline. Note that HCO3− secretion can be directly stimulated by the action of secretagogues on duct cells. The duct epithelium is relatively tight and lacks expression of aquaporin, and therefore water cannot follow the ions rapidly enough to maintain isotonicity at moderate or high flow rates during stimulated salivary secretion. Thus, with an increase in the secretion rate, there is less time for modification by the ducts, and the resulting saliva more closely resembles the primary secretion and therefore plasma. However, [HCO3−] remains high because it is secreted by duct and possibly acinar cells by the action of secretagogues (Fig. 27-2).
< div class='tao-gold-member'>
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


