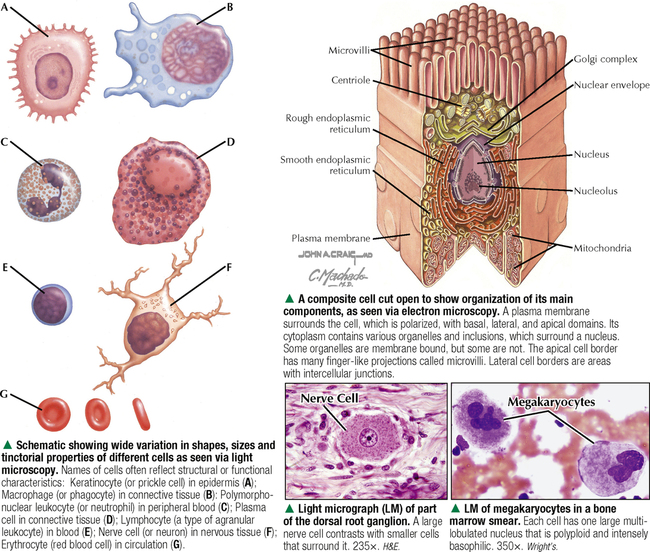
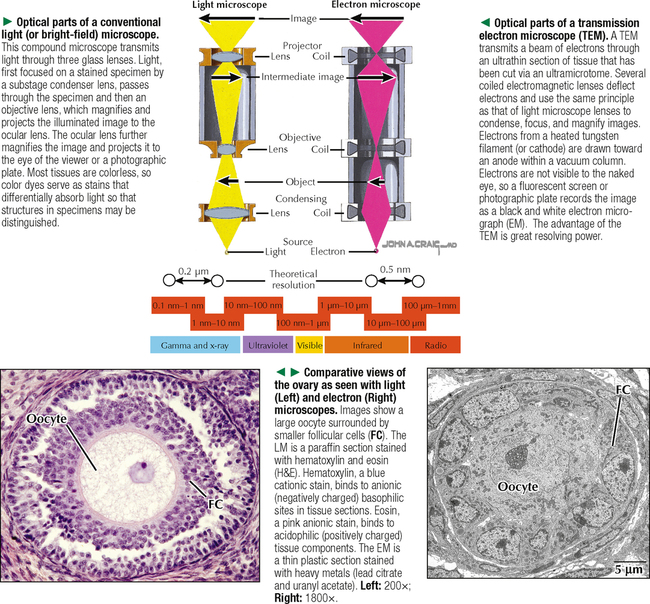
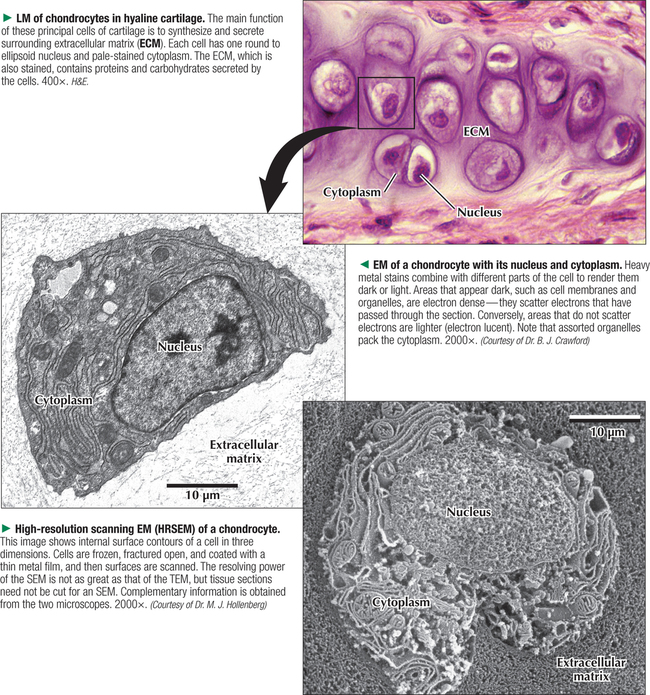
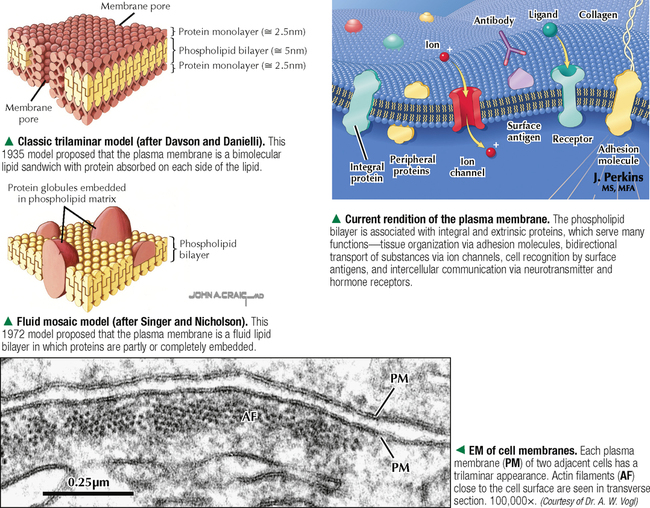
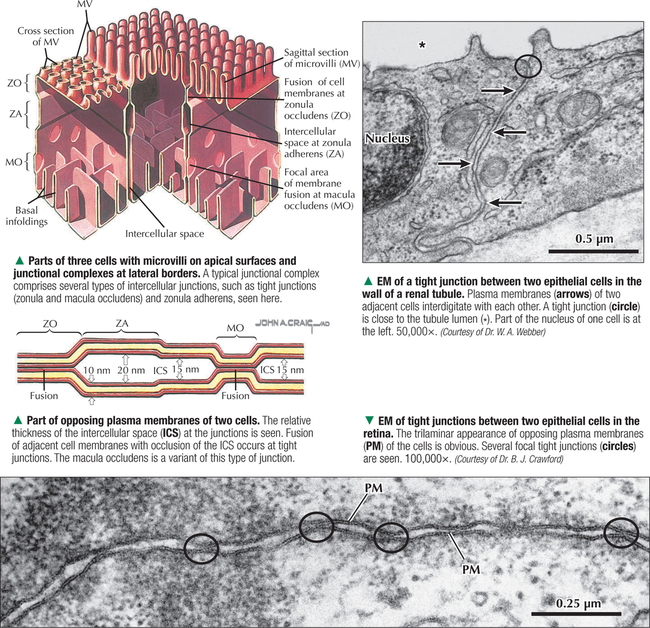
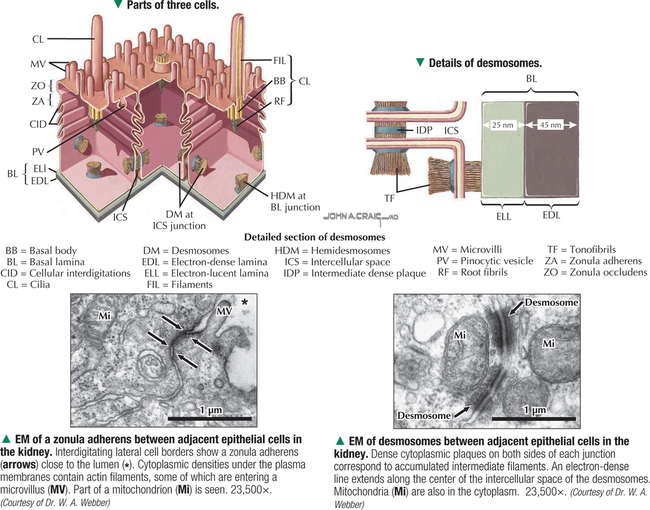
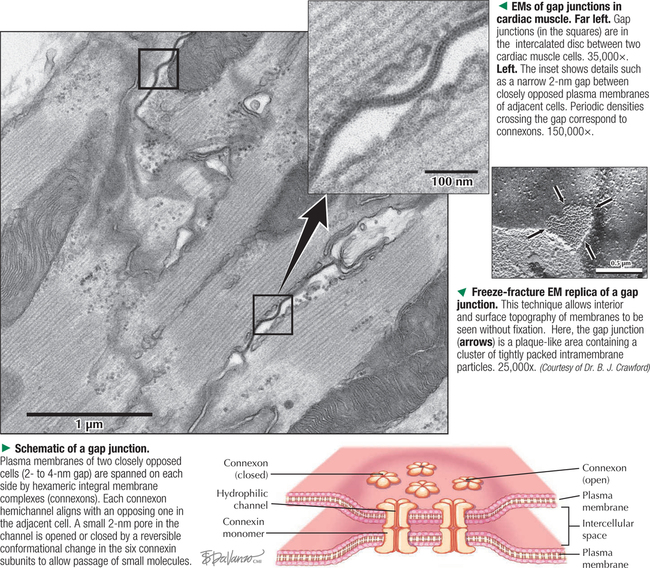
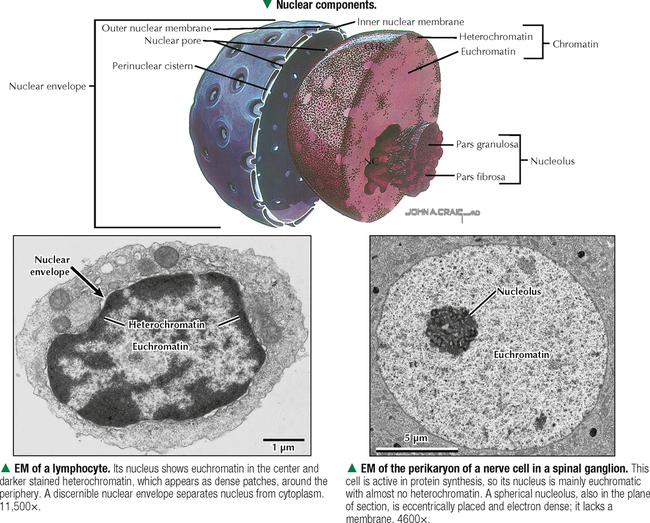
1 THE CELL 1.1. Overview 1.2. Microscopes and Techniques 1.3. Different Appearances of Cells According to Technique 1.4. Ultrastructure and Function of Cell Membranes 1.5. Intercellular Junctions: Ultrastructure and Function of Tight Junctions 1.6. Intercellular Junctions: Ultrastructure and Function of Anchoring Junctions 1.7. Intercellular Junctions: Ultrastructure and Function of Gap Junctions 1.8. Ultrastructure and Function of the Nucleus and Nucleolus 1.9. Ultrastructure and Function of the Nucleus: Chromatin and Matrix 1.10. Ultrastructure and Function of the Nuclear Envelope 1.11. Ultrastructure and Function of Mitochondria 1.12. Ultrastructure and Function of Mitochondrial Cristae and Matrix 1.13. Ultrastructure and Function of Smooth Endoplasmic Reticulum 1.14. Ultrastructure and Function of Rough Endoplasmic Reticulum 1.15. Ultrastructure and Function of Ribosomes 1.16. Ultrastructure of the Golgi Complex 1.17. Functions of the Golgi Complex 1.18. Ultrastructure and Function of Lysosomes 1.19. Ultrastructure and Function of Peroxisomes 1.20. Ultrastructure and Function of Inclusions: Glycogen 1.21. Ultrastructure and Function of Inclusions: Lipid Droplets 1.22. Ultrastructure and Function of Cytoplasmic Vesicles: Endocytosis, Transcytosis, and Exocytosis 1.23. Ultrastructure and Function of Microtubules 1.24. Ultrastructure and Function of Cytoplasmic Filaments 1.25. Ultrastructure and Function of the Centrosome and Centrioles 1.26. The Cell Cycle, Mitosis, and Other Cellular Processes 1.27. Specializations of the Cell Surface: Cilia and Basal Bodies 1.1 OVERVIEW The human body is organized into four basic tissues (epithelial, muscle, nervous, and connective) that consist of cells and associated extracellular matrix. The cell is the fundamental structural and functional unit of all living organisms. The body contains about 60 × 1012 cells—some 200 different types whose size and shape vary widely—but all have a common structural plan. The eukaryotic cell is a mass of protoplasm surrounded by an external plasma (limiting) membrane. The two components of the protoplasm are the nucleus, which holds the genome consisting of chromosomes, and the cytoplasm, a complex aqueous gel made of water (about 70%), proteins, lipids, carbohydrates, and organic and inorganic molecules. Organelles (specialized structures with functional capability) and inclusions (relatively inert, transitory structures) are in the cytoplasm. Except for mature erythrocytes, without a nucleus, most cells have one nucleus that conforms to the cell’s shape. A few cells, such as osteoclasts and skeletal muscle cells, may be multinucleated. A nuclear envelope invests the nucleus, whose substance, called chromatin, contains one or more nucleoli. Internal cell structure is modified to reflect function: Muscle cells, for example, are modified for contraction; nerve cells (or neurons), for conduction; connective tissue cells such as fibroblasts, for support; and glandular epithelial cells, for secretion. HISTORICAL POINT German scientists—biologist Theodor Schwann (1810–1882) and botanist Matthias Schleiden (1804–1881)—proposed the cell theory, which states that all living organisms are composed of similar units of organization called cells. For his observations on normal animal cells, Schwann is recognized as the father of modern histology. Later, renowned German pathologist Rudolph Virchow (1821–1902) proposed that disease originates in cells, not in tissues or organs. Because he was the first to use microscopes and histologic specimens as a basis for the study of pathology, he is credited as the founder of modern cytopathology. With advances in medical science more than a century later, knowing the light and electron microscopic appearance of cells has become fundamental to diagnosis, treatment, and clinical management of many common and rare diseases. 1.2 MICROSCOPES AND TECHNIQUES Histology is the study of body tissues and cells, their constituents. Cells cannot be seen with the naked eye, so the primary tool used to study them is the microscope. It produces enlarged images of cells and enhances contrast for resolving details. Of several kinds of microscopes, two major ones are light and electron microscopes. They have different lenses and sources of illumination and provide complementary information at different levels of resolution and magnification. The ability to discriminate two points that are close together is the resolving power of a microscope. It is related to the light wavelength. A conventional light microscope uses bright-field illumination, with a resolving power of about 0.2 μm. Study specimens absorb visible light; glass lenses focus and magnify specimens. Most cells absorb very little light, so staining is needed to increase light absorption. Cells and tissues first undergo sequential processing steps. Fixation in aldehydes and dehydration in alcohols are followed by embedding in paraffin or plastic. Specimen sections (or slices) are made with a microtome, followed by staining with color dyes. The illumination source of the transmission electron microscope (TEM) is a beam of electrons, which has a smaller wavelength. The resolving power of the TEM, 0.2–0.5 nm, is about 103 greater than that of the light microscope. For the TEM, ultrathin sections are cut after specimens have been fixed and embedded in plastic. Sections are then stained with heavy metals to enhance contrast, and black-and-white, not color, images result. A scanning electron microscope (SEM) is used for thick specimens or whole cells that have been fixed, dried, and coated with a thin metal film. It provides three-dimensional surface views. A high-resolution SEM (HRSEM) allows internal morphology of cells and organelles to be discerned with great depth of focus. 1.3 DIFFERENT APPEARANCES OF CELLS ACCORDING TO TECHNIQUE Histologic techniques provide different but complementary views of cells and thus a useful morphologic base, which can aid understanding of cell function in health and disease. Paraffin sections are routinely stained with hematoxylin and eosin (H&E) and examined with a light microscope. Cell nuclei (which are rich in nucleic acids such as DNA and RNA) have an affinity for hematoxylin (a basic dye), stain blue, and are termed basophilic. In contrast, the cytoplasm of cells and extracellular matrix typically have an affinity for eosin (an anionic dye), stain pink, and are eosinophilic (or acidophilic). With superior resolving power, a TEM provides better elucidation of cell details, such as membranes and organelles, than a light microscope. Different parts of cells have distinct affinities for metal stains used on thin sections, so resulting two-dimensional images show variations in electron density, recorded in black and white. HRSEM images of freeze-fractured cells show three-dimensional spatial relationships of organelles and inclusions. 1.4 ULTRASTRUCTURE AND FUNCTION OF CELL MEMBRANES Membranes—semipermeable barriers that selectively regulate movement of ions, water, and macromolecules—are ubiquitous in cells. They vary in composition depending on cell type and location, but all consist of about 35% lipids, 60% proteins, and 5% carbohydrates. The cell (or plasma) membrane forms an external boundary. Intracellular membranes surround nuclei and membrane-bound organelles. Membranes are beyond the limit of resolution of a light microscope and are thus difficult to visualize without special techniques. By high-magnification electron microscopy, membranes have a trilaminar appearance: two dark lines separated by a thin electron-lucent zone. The entire trilaminar membrane, or unit membrane, is 5–8 nm thick. Membranes are made of a lipid bilayer, with a structure consistent with a highly dynamic fluid mosaic model: two hydrophilic phospholipid leaflets with polar phosphate heads that point outward. The hydrophobic fatty acid tail regions form the internal membrane framework. Cholesterol molecules, dispersed throughout the membrane, impart fluidity to it. Intrinsic (integral) globular proteins lie in the lipid bilayer and span the membrane thickness. Extrinsic (peripheral) proteins are also anchored to the membrane and associate with outside or inside surfaces of the bilayer. Carbohydrates often form a fuzzy coat called the glycocalyx on the outside of membranes. Membranes contain channels and ion pumps made of proteins that regulate the cell’s internal milieu by creating electrical charge differences. Membranes also contain receptors for hormones and growth factors, such as receptors for neurotransmitters in plasma membranes of neurons and muscle cells. CLINICAL POINT Electron microscopy (EM) is indispensable for accurate diagnosis of diseases in which pathologic changes are too small to be resolved by light microscopy (e.g., glomerular basement membrane variations in kidney diseases, poorly differentiated adenocarcinomas, mitochondrial alterations in myopathies, and some skin cancers). In diagnostic virology, EM has contributed to discovery of many clinically important viruses. The differential diagnosis of smallpox (variola) and chickenpox (varicella-zoster) viruses and the discovery of hepatitis B virus were first done by EM using negative staining techniques. More recently, EM has been essential in detecting Ebola, Norwalk, and severe acute respiratory syndrome (SARS) viruses. Also, EM has been instrumental in elucidating mechanisms of virus–host cell interactions in human immunodeficiency virus (HIV) infection, resulting in development of new highly active antiretroviral therapy (HAART) and vaccine strategies. 1.5 INTERCELLULAR JUNCTIONS: ULTRASTRUCTURE AND FUNCTION OF TIGHT JUNCTIONS To increase adhesiveness, most adjacent cells have simple interdigitations between them. Cell membranes interact with extracellular matrix by adhesive contacts consisting of cell adhesion molecules. Cells also show more specialized modifications of plasma membranes—intercellular junctions of different kinds. There are three major types: tight (zonula and macula occludens), anchoring (macula and zonula adherens), and gap (or communicating) junctions. Tight junctions are common between epithelial cells and are closest to the luminal surface, where they form an occluding, belt-like seal between cells. At different sites, they form permeability barriers to prevent indiscriminate passage of material. The tightness and permeability features of these junctions depend on cell type and location. In endothelia of specialized capillaries, they are the basis for the blood-brain, blood-ocular, and blood-testis barriers. In other sites, they define a boundary between apical and basolateral domains of plasma membrane. In high-magnification electron micrographs (EMs), plasma membranes of adjacent cells appear fused at one or more focal contact sites that eliminate intervening extracellular spaces. Each contact site contains transmembrane proteins, such as occludin, and different classes of claudins. Other cytoplasmic proteins, as well as cadherin proteins, reinforce the sites. A freeze-fracture EM shows tight junctions with a network of ridges and opposing grooves, which correspond to transmembrane proteins. Actin filaments of the cytoskeleton also associate with cytoplasmic sides of tight junctions. 1.6 INTERCELLULAR JUNCTIONS: ULTRASTRUCTURE AND FUNCTION OF ANCHORING JUNCTIONS Two kinds of anchoring junctions, zonula adherens and macula adherens (desmosome), hold cells together. They usually occur between lateral borders of adjacent epithelial cells. They resist mechanical stress and prevent lateral disruption by stabilizing the epithelium. Cytoplasmic actin filaments anchor zonulae adherentes; intermediate filaments (tonofilaments) anchor desmosomes. In most epithelia, a zonula adherens usually encircles the apical part of the whole cell just below the tight junction. Transmembrane proteins, consisting mostly of cadherin molecules, are on both sides of the junction. Their extracellular domains span the narrow gap (20 nm) between adjacent cells; their intracellular domains interact with other cytoplasmic proteins (vinculin and α-actinin) to anchor actin filaments of the cytoskeleton. Desmosomes are more complex, plaque-like junctions in epithelial cells, as well as in cardiac and smooth muscle cells, that resemble spot welds and strongly hold cells together at focal points. Dense cytoplasmic plaques are on the cytoplasmic sides of opposing plasma membranes. The intercellular space (20–25 nm wide) often shows a dense line in the center that parallels opposing cell membranes. This space contains transmembrane cadherins (desmogelins and desmocollins) that span it and link adjacent plasma membranes. Accessory proteins in the dense plaques (desmoplakin and plakoglobin) anchor intermediate filaments. Depending on location, desmosomes may have different types of intermediate filaments, such as keratins, associated with epithelial cells, and desmin, in cardiac muscle cells. CLINICAL POINT Claudins, a family of 24 integral membrane proteins, regulate tight junction permeability and epithelial cell polarity. Most types of cancers (known as carcinomas) originate from epithelial cells; alterations in claudin expression contribute to initiation of many such malignancies (e.g., metastatic breast cancer, colorectal carcinoma, mesothelioma, prostate cancer) via epithelial barrier breakdown. Because claudin expression appears to be specific for different kinds and stages of tumors, such knowledge may be useful in confirming histologic diagnosis, predicting prognosis, and serving as targets in cancer therapy. Also, diminished expression of E-cadherins—a class of transmembrane proteins localized in desmosomes—contributes to certain forms of breast, endometrial, and ovarian cancers whereby concomitant loss of cell adhesion correlates with increased cancer invasiveness and metastasis. Developing novel methods to block E-cadherin downregulation may be useful in future approaches to gene therapy. 1.7 INTERCELLULAR JUNCTIONS: ULTRASTRUCTURE AND FUNCTION OF GAP JUNCTIONS Metabolic, ionic, and low-resistance electrical communication occurs between adjacent cells via gap junctions, in which a narrow gap of about 2 nm separates opposing cell membranes. Gap junctions are difficult to discern in routine EMs; elucidation of structural details requires freeze-fracture methods or other techniques that use immunocytochemistry with antibody probes. Gap junctions are specialized sites composed of large, tightly packed intercellular channels, which connect cytoplasm of adjacent cells. Each cylindrical channel, 10–12 nm long and 2.8–3.0 nm in diameter, consists of a pair of half-channels, termed connexons, which are embedded in the cell membranes. Each connexon comprises six symmetric protein subunits, called connexins, that are transmembrane proteins surrounding a small central aqueous pore (diameter: 1.5–2.0 nm). Across the narrow gap, a connexon of one cell is aligned with that of the adjacent cell so that the central pores form one continuous conduit, thereby allowing direct communication. No leakage of ions or fluid out of cells and into the extracellular space occurs. Each of about 20 different connexin proteins has a separate gene encoding it. Connexons, like other voltage-gated channels in membranes, can undergo reversible conformational changes to open or close gap junction channels. CLINICAL POINT Several diseases result from mutations in genes encoding connexins, which are named according to molecular size. Recessive mutations in connexin-26, with a molecular size of 26 kD, lead to the most common cause of inherited human deafness, which often affects the elderly. Connexin-26 is usually involved in K+ transport in cells that support cochlear hair cells. An X-linked form of Charcot-Marie-Tooth disease is due to mutations in connexin-32 and causes degeneration of myelin sheaths in central and peripheral nervous systems. A mutation in connexin-50 leads to cataracts in the lens of the eye. 1.8 ULTRASTRUCTURE AND FUNCTION OF THE NUCLEUS AND NUCLEOLUS The nucleus—the largest, most conspicuous structure in the cell—contains genetic material. Size and shape may depend on cell type: Usually spherical or ellipsoidal, a nucleus may also be elongated (as in columnar epithelial cells) or lobulated (as in polymorphonuclear leukocytes and megakaryocytes). Most cells have one nucleus; some (e.g., hepatocytes) may be binucleated, others (e.g., osteoclasts, skeletal muscle fibers), multinucleated. The nucleus consists of nucleolus, chromatin, nuclear matrix, and nuclear envelope. The nucleolus, the most conspicuous part of the nucleus, is a dense, ovoid, discrete area (up to 1 μm in diameter) with no membrane around it. Its size, number, and location may depend on a cell’s functional activity. The nucleolus is the site of ribosomal RNA (rRNA) transcription and production of ribosomes. It has a high content of RNA, so it is intensely basophilic by light microscopy. In EMs, the nucleolus shows two areas, the pars granulosa and pars fibrosa, that have no clear boundary between them. The pars granulosa, in peripheral nucleolar regions, is the main site of preribosome assembly. It consists of densely packed clusters of preribosomal particles (diameter: 15–20 nm) that are rich in ribonucleoprotein. The more central pars fibrosa contains a dense, irregular network of fine filaments (5 nm in diameter), rRNA genes, and transcription factors. The nucleolus disassembles in the prophase of mitosis but then reorganizes in daughter cells when cell division is complete. CLINICAL POINT Structural and functional abnormalities in nucleoli have been implicated in the progression of many diseases, including several forms of cancer. Tumor cells in high-grade malignancies, such as carcinomas of the breast, liver, and prostate, commonly have multiple nucleoli that are larger and more irregular in shape than in normal cells. Nucleoli normally contain proteins for ribosome biosynthesis and nucleolar oncogenes and tumor suppressors that may control cancer cell proliferation. Nucleolar organizing regions (NORs) are vital segments in the nucleus around which nucleoli reform during telophase. NORs contain argyrophilic proteins, such as nucleolin and nucleophosmin Only gold members can continue reading. Log In or Register to continue Share this: Share on X (Opens in new window) X Share on Facebook (Opens in new window) Facebook Like this:Like Loading… Related Related posts: CARDIOVASCULAR SYSTEM RESPIRATORY SYSTEM SPECIAL SENSES FEMALE REPRODUCTIVE SYSTEM Stay updated, free articles. Join our Telegram channel Join Tags: Netters Essential Histology Jun 18, 2016 | Posted by admin in HISTOLOGY | Comments Off on THE CELL Full access? Get Clinical Tree
1 THE CELL 1.1. Overview 1.2. Microscopes and Techniques 1.3. Different Appearances of Cells According to Technique 1.4. Ultrastructure and Function of Cell Membranes 1.5. Intercellular Junctions: Ultrastructure and Function of Tight Junctions 1.6. Intercellular Junctions: Ultrastructure and Function of Anchoring Junctions 1.7. Intercellular Junctions: Ultrastructure and Function of Gap Junctions 1.8. Ultrastructure and Function of the Nucleus and Nucleolus 1.9. Ultrastructure and Function of the Nucleus: Chromatin and Matrix 1.10. Ultrastructure and Function of the Nuclear Envelope 1.11. Ultrastructure and Function of Mitochondria 1.12. Ultrastructure and Function of Mitochondrial Cristae and Matrix 1.13. Ultrastructure and Function of Smooth Endoplasmic Reticulum 1.14. Ultrastructure and Function of Rough Endoplasmic Reticulum 1.15. Ultrastructure and Function of Ribosomes 1.16. Ultrastructure of the Golgi Complex 1.17. Functions of the Golgi Complex 1.18. Ultrastructure and Function of Lysosomes 1.19. Ultrastructure and Function of Peroxisomes 1.20. Ultrastructure and Function of Inclusions: Glycogen 1.21. Ultrastructure and Function of Inclusions: Lipid Droplets 1.22. Ultrastructure and Function of Cytoplasmic Vesicles: Endocytosis, Transcytosis, and Exocytosis 1.23. Ultrastructure and Function of Microtubules 1.24. Ultrastructure and Function of Cytoplasmic Filaments 1.25. Ultrastructure and Function of the Centrosome and Centrioles 1.26. The Cell Cycle, Mitosis, and Other Cellular Processes 1.27. Specializations of the Cell Surface: Cilia and Basal Bodies 1.1 OVERVIEW The human body is organized into four basic tissues (epithelial, muscle, nervous, and connective) that consist of cells and associated extracellular matrix. The cell is the fundamental structural and functional unit of all living organisms. The body contains about 60 × 1012 cells—some 200 different types whose size and shape vary widely—but all have a common structural plan. The eukaryotic cell is a mass of protoplasm surrounded by an external plasma (limiting) membrane. The two components of the protoplasm are the nucleus, which holds the genome consisting of chromosomes, and the cytoplasm, a complex aqueous gel made of water (about 70%), proteins, lipids, carbohydrates, and organic and inorganic molecules. Organelles (specialized structures with functional capability) and inclusions (relatively inert, transitory structures) are in the cytoplasm. Except for mature erythrocytes, without a nucleus, most cells have one nucleus that conforms to the cell’s shape. A few cells, such as osteoclasts and skeletal muscle cells, may be multinucleated. A nuclear envelope invests the nucleus, whose substance, called chromatin, contains one or more nucleoli. Internal cell structure is modified to reflect function: Muscle cells, for example, are modified for contraction; nerve cells (or neurons), for conduction; connective tissue cells such as fibroblasts, for support; and glandular epithelial cells, for secretion. HISTORICAL POINT German scientists—biologist Theodor Schwann (1810–1882) and botanist Matthias Schleiden (1804–1881)—proposed the cell theory, which states that all living organisms are composed of similar units of organization called cells. For his observations on normal animal cells, Schwann is recognized as the father of modern histology. Later, renowned German pathologist Rudolph Virchow (1821–1902) proposed that disease originates in cells, not in tissues or organs. Because he was the first to use microscopes and histologic specimens as a basis for the study of pathology, he is credited as the founder of modern cytopathology. With advances in medical science more than a century later, knowing the light and electron microscopic appearance of cells has become fundamental to diagnosis, treatment, and clinical management of many common and rare diseases. 1.2 MICROSCOPES AND TECHNIQUES Histology is the study of body tissues and cells, their constituents. Cells cannot be seen with the naked eye, so the primary tool used to study them is the microscope. It produces enlarged images of cells and enhances contrast for resolving details. Of several kinds of microscopes, two major ones are light and electron microscopes. They have different lenses and sources of illumination and provide complementary information at different levels of resolution and magnification. The ability to discriminate two points that are close together is the resolving power of a microscope. It is related to the light wavelength. A conventional light microscope uses bright-field illumination, with a resolving power of about 0.2 μm. Study specimens absorb visible light; glass lenses focus and magnify specimens. Most cells absorb very little light, so staining is needed to increase light absorption. Cells and tissues first undergo sequential processing steps. Fixation in aldehydes and dehydration in alcohols are followed by embedding in paraffin or plastic. Specimen sections (or slices) are made with a microtome, followed by staining with color dyes. The illumination source of the transmission electron microscope (TEM) is a beam of electrons, which has a smaller wavelength. The resolving power of the TEM, 0.2–0.5 nm, is about 103 greater than that of the light microscope. For the TEM, ultrathin sections are cut after specimens have been fixed and embedded in plastic. Sections are then stained with heavy metals to enhance contrast, and black-and-white, not color, images result. A scanning electron microscope (SEM) is used for thick specimens or whole cells that have been fixed, dried, and coated with a thin metal film. It provides three-dimensional surface views. A high-resolution SEM (HRSEM) allows internal morphology of cells and organelles to be discerned with great depth of focus. 1.3 DIFFERENT APPEARANCES OF CELLS ACCORDING TO TECHNIQUE Histologic techniques provide different but complementary views of cells and thus a useful morphologic base, which can aid understanding of cell function in health and disease. Paraffin sections are routinely stained with hematoxylin and eosin (H&E) and examined with a light microscope. Cell nuclei (which are rich in nucleic acids such as DNA and RNA) have an affinity for hematoxylin (a basic dye), stain blue, and are termed basophilic. In contrast, the cytoplasm of cells and extracellular matrix typically have an affinity for eosin (an anionic dye), stain pink, and are eosinophilic (or acidophilic). With superior resolving power, a TEM provides better elucidation of cell details, such as membranes and organelles, than a light microscope. Different parts of cells have distinct affinities for metal stains used on thin sections, so resulting two-dimensional images show variations in electron density, recorded in black and white. HRSEM images of freeze-fractured cells show three-dimensional spatial relationships of organelles and inclusions. 1.4 ULTRASTRUCTURE AND FUNCTION OF CELL MEMBRANES Membranes—semipermeable barriers that selectively regulate movement of ions, water, and macromolecules—are ubiquitous in cells. They vary in composition depending on cell type and location, but all consist of about 35% lipids, 60% proteins, and 5% carbohydrates. The cell (or plasma) membrane forms an external boundary. Intracellular membranes surround nuclei and membrane-bound organelles. Membranes are beyond the limit of resolution of a light microscope and are thus difficult to visualize without special techniques. By high-magnification electron microscopy, membranes have a trilaminar appearance: two dark lines separated by a thin electron-lucent zone. The entire trilaminar membrane, or unit membrane, is 5–8 nm thick. Membranes are made of a lipid bilayer, with a structure consistent with a highly dynamic fluid mosaic model: two hydrophilic phospholipid leaflets with polar phosphate heads that point outward. The hydrophobic fatty acid tail regions form the internal membrane framework. Cholesterol molecules, dispersed throughout the membrane, impart fluidity to it. Intrinsic (integral) globular proteins lie in the lipid bilayer and span the membrane thickness. Extrinsic (peripheral) proteins are also anchored to the membrane and associate with outside or inside surfaces of the bilayer. Carbohydrates often form a fuzzy coat called the glycocalyx on the outside of membranes. Membranes contain channels and ion pumps made of proteins that regulate the cell’s internal milieu by creating electrical charge differences. Membranes also contain receptors for hormones and growth factors, such as receptors for neurotransmitters in plasma membranes of neurons and muscle cells. CLINICAL POINT Electron microscopy (EM) is indispensable for accurate diagnosis of diseases in which pathologic changes are too small to be resolved by light microscopy (e.g., glomerular basement membrane variations in kidney diseases, poorly differentiated adenocarcinomas, mitochondrial alterations in myopathies, and some skin cancers). In diagnostic virology, EM has contributed to discovery of many clinically important viruses. The differential diagnosis of smallpox (variola) and chickenpox (varicella-zoster) viruses and the discovery of hepatitis B virus were first done by EM using negative staining techniques. More recently, EM has been essential in detecting Ebola, Norwalk, and severe acute respiratory syndrome (SARS) viruses. Also, EM has been instrumental in elucidating mechanisms of virus–host cell interactions in human immunodeficiency virus (HIV) infection, resulting in development of new highly active antiretroviral therapy (HAART) and vaccine strategies. 1.5 INTERCELLULAR JUNCTIONS: ULTRASTRUCTURE AND FUNCTION OF TIGHT JUNCTIONS To increase adhesiveness, most adjacent cells have simple interdigitations between them. Cell membranes interact with extracellular matrix by adhesive contacts consisting of cell adhesion molecules. Cells also show more specialized modifications of plasma membranes—intercellular junctions of different kinds. There are three major types: tight (zonula and macula occludens), anchoring (macula and zonula adherens), and gap (or communicating) junctions. Tight junctions are common between epithelial cells and are closest to the luminal surface, where they form an occluding, belt-like seal between cells. At different sites, they form permeability barriers to prevent indiscriminate passage of material. The tightness and permeability features of these junctions depend on cell type and location. In endothelia of specialized capillaries, they are the basis for the blood-brain, blood-ocular, and blood-testis barriers. In other sites, they define a boundary between apical and basolateral domains of plasma membrane. In high-magnification electron micrographs (EMs), plasma membranes of adjacent cells appear fused at one or more focal contact sites that eliminate intervening extracellular spaces. Each contact site contains transmembrane proteins, such as occludin, and different classes of claudins. Other cytoplasmic proteins, as well as cadherin proteins, reinforce the sites. A freeze-fracture EM shows tight junctions with a network of ridges and opposing grooves, which correspond to transmembrane proteins. Actin filaments of the cytoskeleton also associate with cytoplasmic sides of tight junctions. 1.6 INTERCELLULAR JUNCTIONS: ULTRASTRUCTURE AND FUNCTION OF ANCHORING JUNCTIONS Two kinds of anchoring junctions, zonula adherens and macula adherens (desmosome), hold cells together. They usually occur between lateral borders of adjacent epithelial cells. They resist mechanical stress and prevent lateral disruption by stabilizing the epithelium. Cytoplasmic actin filaments anchor zonulae adherentes; intermediate filaments (tonofilaments) anchor desmosomes. In most epithelia, a zonula adherens usually encircles the apical part of the whole cell just below the tight junction. Transmembrane proteins, consisting mostly of cadherin molecules, are on both sides of the junction. Their extracellular domains span the narrow gap (20 nm) between adjacent cells; their intracellular domains interact with other cytoplasmic proteins (vinculin and α-actinin) to anchor actin filaments of the cytoskeleton. Desmosomes are more complex, plaque-like junctions in epithelial cells, as well as in cardiac and smooth muscle cells, that resemble spot welds and strongly hold cells together at focal points. Dense cytoplasmic plaques are on the cytoplasmic sides of opposing plasma membranes. The intercellular space (20–25 nm wide) often shows a dense line in the center that parallels opposing cell membranes. This space contains transmembrane cadherins (desmogelins and desmocollins) that span it and link adjacent plasma membranes. Accessory proteins in the dense plaques (desmoplakin and plakoglobin) anchor intermediate filaments. Depending on location, desmosomes may have different types of intermediate filaments, such as keratins, associated with epithelial cells, and desmin, in cardiac muscle cells. CLINICAL POINT Claudins, a family of 24 integral membrane proteins, regulate tight junction permeability and epithelial cell polarity. Most types of cancers (known as carcinomas) originate from epithelial cells; alterations in claudin expression contribute to initiation of many such malignancies (e.g., metastatic breast cancer, colorectal carcinoma, mesothelioma, prostate cancer) via epithelial barrier breakdown. Because claudin expression appears to be specific for different kinds and stages of tumors, such knowledge may be useful in confirming histologic diagnosis, predicting prognosis, and serving as targets in cancer therapy. Also, diminished expression of E-cadherins—a class of transmembrane proteins localized in desmosomes—contributes to certain forms of breast, endometrial, and ovarian cancers whereby concomitant loss of cell adhesion correlates with increased cancer invasiveness and metastasis. Developing novel methods to block E-cadherin downregulation may be useful in future approaches to gene therapy. 1.7 INTERCELLULAR JUNCTIONS: ULTRASTRUCTURE AND FUNCTION OF GAP JUNCTIONS Metabolic, ionic, and low-resistance electrical communication occurs between adjacent cells via gap junctions, in which a narrow gap of about 2 nm separates opposing cell membranes. Gap junctions are difficult to discern in routine EMs; elucidation of structural details requires freeze-fracture methods or other techniques that use immunocytochemistry with antibody probes. Gap junctions are specialized sites composed of large, tightly packed intercellular channels, which connect cytoplasm of adjacent cells. Each cylindrical channel, 10–12 nm long and 2.8–3.0 nm in diameter, consists of a pair of half-channels, termed connexons, which are embedded in the cell membranes. Each connexon comprises six symmetric protein subunits, called connexins, that are transmembrane proteins surrounding a small central aqueous pore (diameter: 1.5–2.0 nm). Across the narrow gap, a connexon of one cell is aligned with that of the adjacent cell so that the central pores form one continuous conduit, thereby allowing direct communication. No leakage of ions or fluid out of cells and into the extracellular space occurs. Each of about 20 different connexin proteins has a separate gene encoding it. Connexons, like other voltage-gated channels in membranes, can undergo reversible conformational changes to open or close gap junction channels. CLINICAL POINT Several diseases result from mutations in genes encoding connexins, which are named according to molecular size. Recessive mutations in connexin-26, with a molecular size of 26 kD, lead to the most common cause of inherited human deafness, which often affects the elderly. Connexin-26 is usually involved in K+ transport in cells that support cochlear hair cells. An X-linked form of Charcot-Marie-Tooth disease is due to mutations in connexin-32 and causes degeneration of myelin sheaths in central and peripheral nervous systems. A mutation in connexin-50 leads to cataracts in the lens of the eye. 1.8 ULTRASTRUCTURE AND FUNCTION OF THE NUCLEUS AND NUCLEOLUS The nucleus—the largest, most conspicuous structure in the cell—contains genetic material. Size and shape may depend on cell type: Usually spherical or ellipsoidal, a nucleus may also be elongated (as in columnar epithelial cells) or lobulated (as in polymorphonuclear leukocytes and megakaryocytes). Most cells have one nucleus; some (e.g., hepatocytes) may be binucleated, others (e.g., osteoclasts, skeletal muscle fibers), multinucleated. The nucleus consists of nucleolus, chromatin, nuclear matrix, and nuclear envelope. The nucleolus, the most conspicuous part of the nucleus, is a dense, ovoid, discrete area (up to 1 μm in diameter) with no membrane around it. Its size, number, and location may depend on a cell’s functional activity. The nucleolus is the site of ribosomal RNA (rRNA) transcription and production of ribosomes. It has a high content of RNA, so it is intensely basophilic by light microscopy. In EMs, the nucleolus shows two areas, the pars granulosa and pars fibrosa, that have no clear boundary between them. The pars granulosa, in peripheral nucleolar regions, is the main site of preribosome assembly. It consists of densely packed clusters of preribosomal particles (diameter: 15–20 nm) that are rich in ribonucleoprotein. The more central pars fibrosa contains a dense, irregular network of fine filaments (5 nm in diameter), rRNA genes, and transcription factors. The nucleolus disassembles in the prophase of mitosis but then reorganizes in daughter cells when cell division is complete. CLINICAL POINT Structural and functional abnormalities in nucleoli have been implicated in the progression of many diseases, including several forms of cancer. Tumor cells in high-grade malignancies, such as carcinomas of the breast, liver, and prostate, commonly have multiple nucleoli that are larger and more irregular in shape than in normal cells. Nucleoli normally contain proteins for ribosome biosynthesis and nucleolar oncogenes and tumor suppressors that may control cancer cell proliferation. Nucleolar organizing regions (NORs) are vital segments in the nucleus around which nucleoli reform during telophase. NORs contain argyrophilic proteins, such as nucleolin and nucleophosmin Only gold members can continue reading. Log In or Register to continue Share this: Share on X (Opens in new window) X Share on Facebook (Opens in new window) Facebook Like this:Like Loading… Related Related posts: CARDIOVASCULAR SYSTEM RESPIRATORY SYSTEM SPECIAL SENSES FEMALE REPRODUCTIVE SYSTEM Stay updated, free articles. Join our Telegram channel Join Tags: Netters Essential Histology Jun 18, 2016 | Posted by admin in HISTOLOGY | Comments Off on THE CELL Full access? Get Clinical Tree