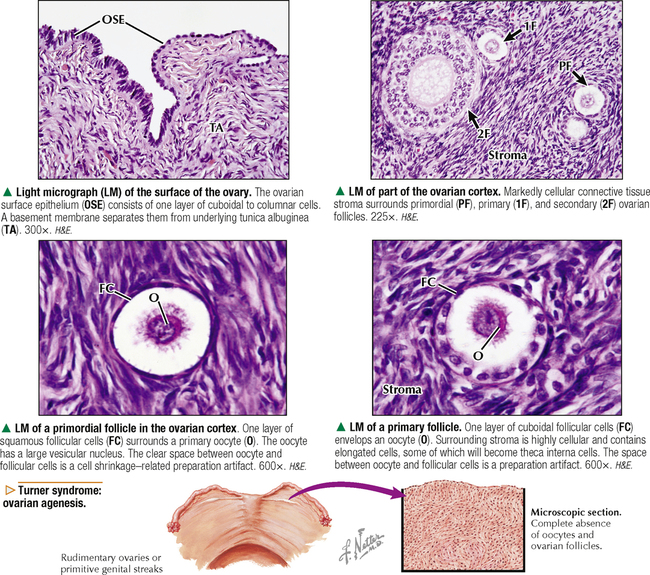
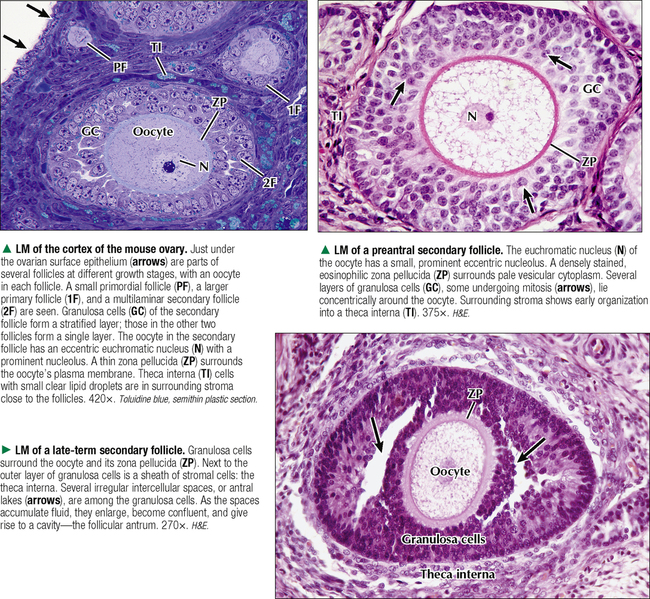
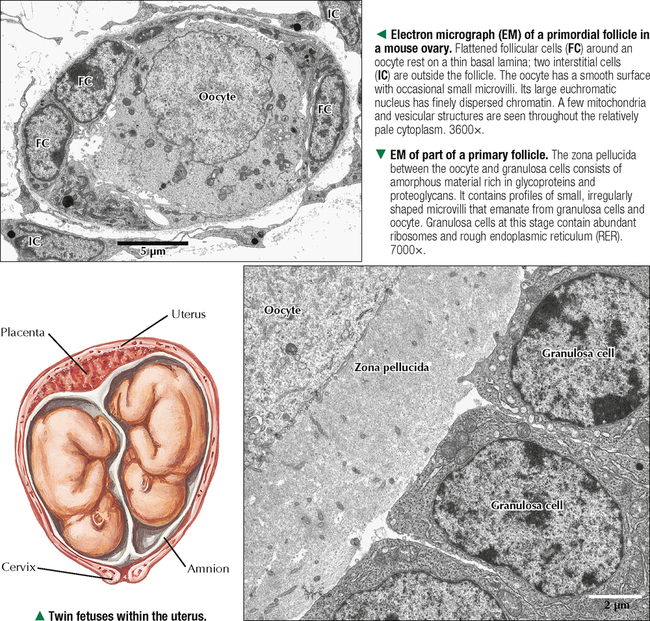
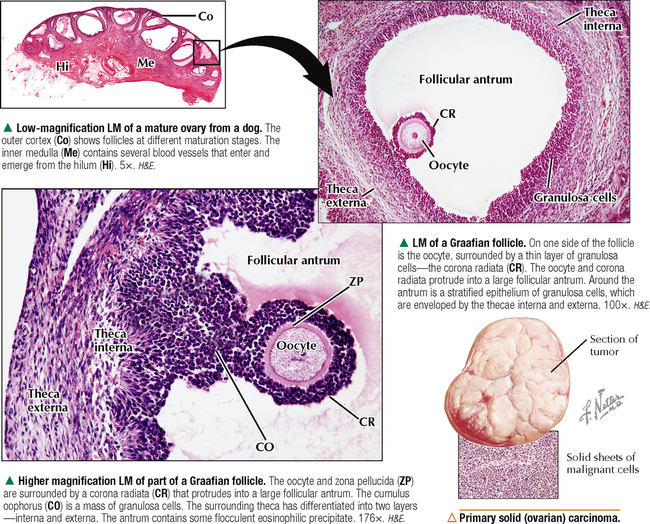
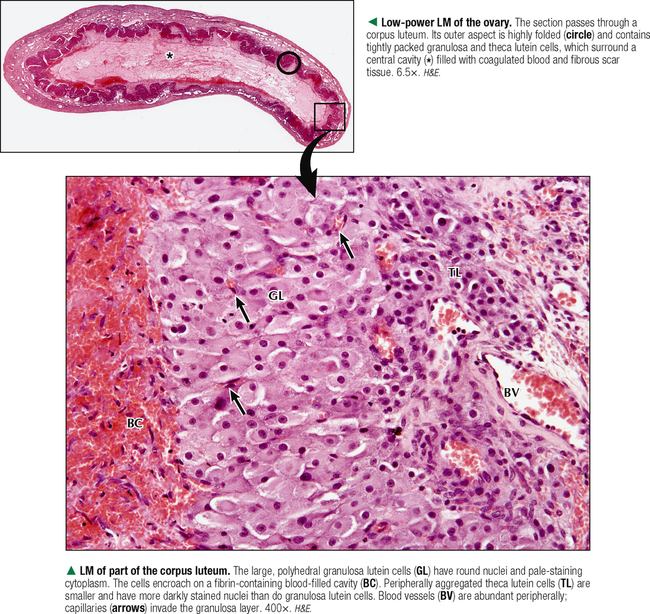
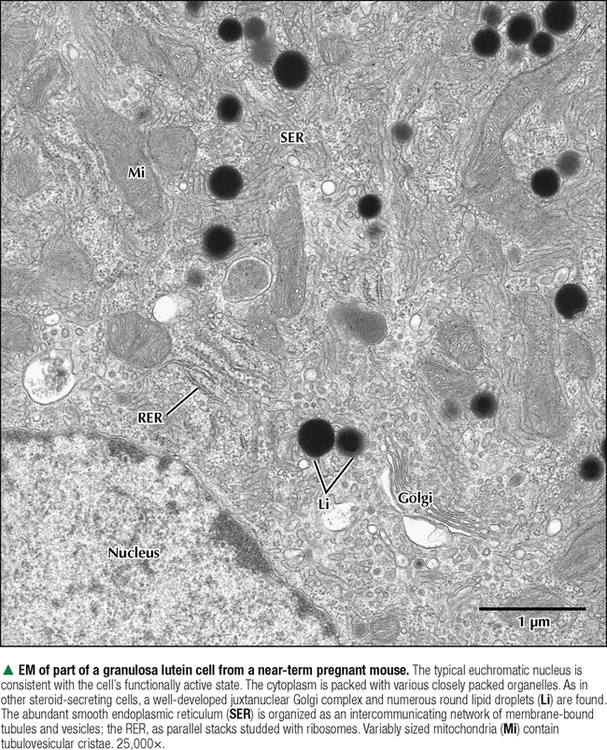
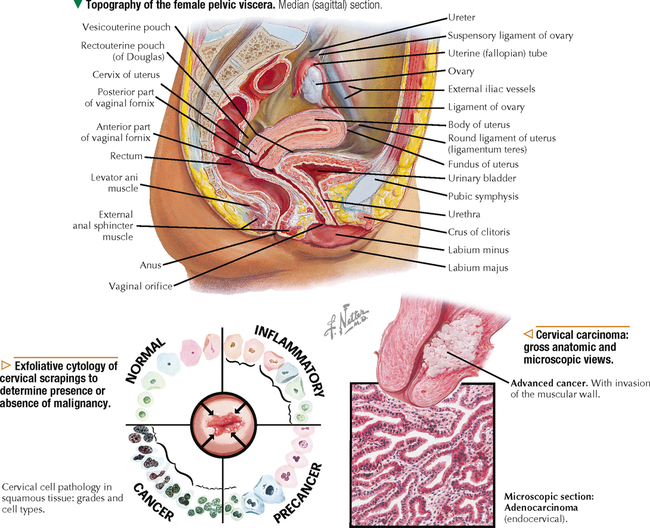
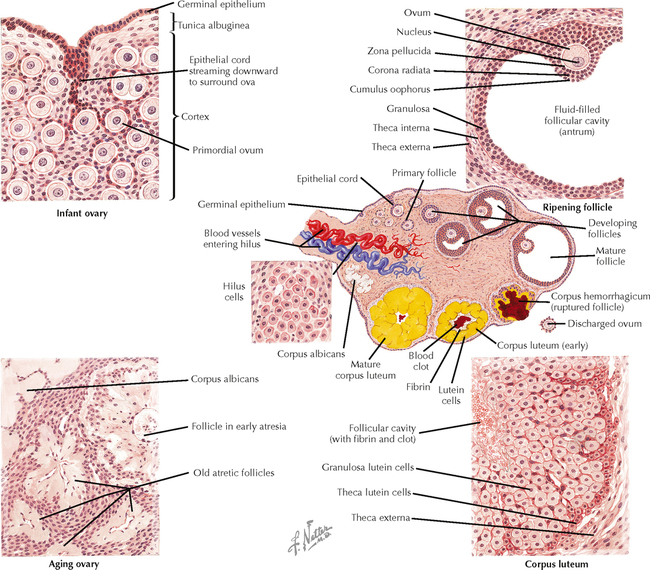
18 The female reproductive system consists of paired ovaries and the genital tract, including fallopian tubes (oviducts, or uterine tubes), uterus, cervix, and vagina, located in the pelvis—the internal genitalia. External genitalia consist of labia majora, labia minora, and clitoris. Mammary glands (see Chapter 2) and placenta are not classified as genital organs but are functionally associated with them. Ovaries, the center of cyclic changes in the female reproductive system, produce female germ cells (ova) and steroid hormones. Fallopian tubes are sites for fertilization of ova, and the uterus harbors fertilized ova during gestation. Like ovaries, the uterus undergoes a regular sequence of changes known as the menstrual cycle. The vagina connects the internal genitalia with the exterior. Embryonic development of the female reproductive system, as in the male, closely parallels that of the urinary system. The system derives mainly from a urogenital ridge of intermediate mesoderm in the posterior abdominal wall. At 6 weeks of gestation, primordial germ cells migrate from their origin in the yolk sac endoderm to the urogenital ridge. Gonad development proceeds with interaction of germ cells with surrounding mesenchyme and coelomic surface epithelium. Germ cells in the primitive ovary develop into oogonia; surface epithelium differentiates into follicular cells. The female genital duct system and external genitalia then develop under the influence of circulating fetal hormones. The paramesonephric (Müllerian) duct system gives rise to most of the genital duct system, and the lower part of the vagina originates from the urogenital sinus.
FEMALE REPRODUCTIVE SYSTEM
18.1 OVERVIEW
Basicmedical Key
Fastest Basicmedical Insight Engine