7
Pathogenesis
CHAPTER CONTENTS
PRINCIPLES OF PATHOGENESIS
A microorganism is a pathogen if it is capable of causing disease; however, some organisms are highly pathogenic (i.e., they often cause disease), whereas others cause disease rarely. Opportunistic pathogens are those that rarely, if ever, cause disease in immunocompetent people but can cause serious infection in immunocompromised patients. These opportunists are frequent members of the body’s normal flora. The origin of the term opportunistic refers to the ability of the organism to take the opportunity offered by reduced host defenses to cause disease.
Virulence is a quantitative measure of pathogenicity and is measured by the number of organisms required to cause disease. The 50% lethal dose (LD50) is the number of organisms needed to kill half the hosts, and the 50% infectious dose (ID50) is the number needed to cause infection in half the hosts. Organisms with a lower LD50 (or ID50) are said to be more virulent than those with a higher LD50 (or ID50) because fewer organisms are needed to cause death or disease.
The infectious dose of an organism required to cause disease varies greatly among the pathogenic bacteria. For example, Shigella and Salmonella both cause diarrhea by infecting the gastrointestinal tract, but the infectious dose of Shigella is less than 100 organisms, whereas the infectious dose of Salmonella is on the order of 100,000 organisms. The infectious dose of bacteria depends primarily on their virulence factors (e.g., whether their pili allow them to adhere well to mucous membranes, whether they produce exotoxins or endotoxins, whether they possess a capsule to protect them from phagocytosis, and whether they can survive various nonspecific host defenses such as acid in the stomach).
There are two uses of the word parasite. Within the context of this chapter, the term refers to the parasitic relationship of the bacteria to the host cells (i.e., the presence of the bacteria is detrimental to the host cells). Bacteria that are human pathogens can be thought of, therefore, as parasites. Some bacterial pathogens are obligate intracellular parasites (e.g., Chlamydia and Rickettsia), because they can grow only within host cells. Many bacteria are facultative parasites because they can grow within cells, outside cells, or on bacteriologic media. The other use of the term parasite refers to the protozoa and the helminths, which are discussed in Part VI of this book.
WHY DO PEOPLE GET INFECTIOUS DISEASES?
People get infectious diseases when microorganisms overpower our host defenses (i.e., when the balance between the organism and the host shifts in favor of the organism). The organism or its products are then present in sufficient amount to induce various symptoms, such as fever and inflammation, which we interpret as those of an infectious disease.
From the organism’s perspective, the two critical determinants in overpowering the host are the number of organisms to which the host, or person, is exposed and the virulence of these organisms. Clearly, the greater the number of organisms, the greater is the likelihood of infection. It is important to realize, however, that a small number of highly virulent organisms can cause disease just as a large number of less virulent organisms can. The virulence of an organism is determined by its ability to produce various virulence factors, several of which were described previously.
The production of specific virulence factors also determines what disease the bacteria cause. For example, a strain of Escherichia coli that produces one type of exotoxin causes watery (nonbloody) diarrhea, whereas a different strain of E. coli that produces another type of exotoxin causes bloody diarrhea. This chapter describes several important examples of specific diseases related to the production of various virulence factors.
From the host’s perspective, the two main arms of our host defenses are innate immunity and acquired immunity, the latter of which includes both antibody-mediated and cell-mediated immunity. A reduction in the functioning of any component of our host defenses shifts the balance in favor of the organism and increases the chance that an infectious disease will occur. Some important causes of a reduction in our host defenses include genetic immunodeficiencies such as agammaglobulinemia and acquired immunodeficiencies such as acquired immunodeficiency syndrome (AIDS), drug-induced immunosuppression in patients with organ transplants, and cancer patients who are receiving chemotherapy. Patients with diabetes and autoimmune diseases also may have reduced host defenses. An overview of our host defenses is presented in Chapters 8 and 57.
In many instances, a person acquires an organism, but no infectious disease occurs because the host defenses were successful. Such asymptomatic infections are very common and are typically recognized by detecting antibody against the organism in the patient’s serum.
TYPES OF BACTERIAL INFECTIONS
The term infection has more than one meaning. One meaning is that an organism has infected the person (i.e., it has entered the body of that person). For example, a person can be infected with an organism of low pathogenicity and not develop symptoms of disease. Another meaning of the term infection is to describe an infectious disease, such as when a person says, “I have an infection.” In this instance, infection and disease are being used interchangeably, but it is important to realize that according to the first definition, the word infection does not have to be equated with disease. Usually, the meaning will be apparent from the context.
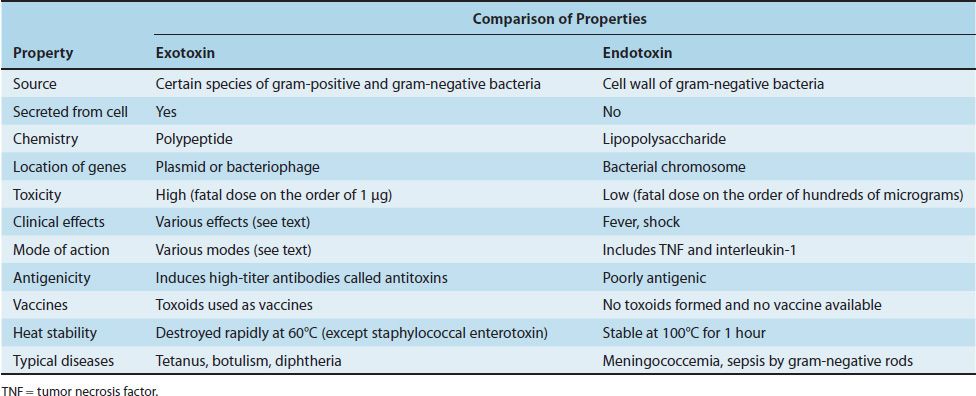
Bacteria cause disease by two major mechanisms: (1) toxin production and (2) invasion and inflammation. Toxins fall into two general categories: exotoxins and endotoxins. Exotoxins are polypeptides released by the cell, whereas endotoxins are lipopolysaccharides (LPS), which form an integral part of the cell wall. Endotoxins occur only in gram-negative rods and cocci, are not actively released from the cell, and cause fever, shock, and other generalized symptoms. Both exotoxins and endotoxins by themselves can cause symptoms; the presence of the bacteria in the host is not required. Invasive bacteria, on the other hand, grow to large numbers locally and induce an inflammatory response consisting of erythema, edema, warmth, and pain. Invasion and inflammation are discussed later in the section entitled “Determinants of Bacterial Pathogenesis.”
Many, but not all, infections are communicable (i.e., they are spread from host to host). For example, tuberculosis is communicable (i.e., it is spread from person to person via airborne droplets produced by coughing), but botulism is not, because the exotoxin produced by the organism in the contaminated food affects only those eating that food. If a disease is highly communicable, the term contagious is applied.
An infection is epidemic if it occurs much more frequently than usual; it is pandemic if it has a worldwide distribution. An endemic infection is constantly present at a low level in a specific population. In addition to infections that result in overt symptoms, many are inapparent or subclinical and can be detected only by demonstrating a rise in antibody titer or by isolating the organism. Some infections result in a latent state, after which reactivation of the growth of the organism and recurrence of symptoms may occur. Certain other infections lead to a chronic carrier state, in which the organisms continue to grow with or without producing symptoms in the host. Chronic carriers (e.g., “Typhoid Mary”) are an important source of infection of others and hence are a public health hazard.
The determination of whether an organism recovered from a patient is actually the cause of the disease involves an awareness of two phenomena: normal flora and colonization. Members of the normal flora are permanent residents of the body and vary in type according to anatomic site (see Chapter 6). When an organism is obtained from a patient’s specimen, the question of whether it is a member of the normal flora is important in interpreting the finding. Colonization refers to the presence of a new organism that is neither a member of the normal flora nor the cause of symptoms. It can be a difficult clinical dilemma to distinguish between a pathogen and a colonizer, especially in specimens obtained from the respiratory tract, such as throat cultures and sputum cultures.
STAGES OF BACTERIAL PATHOGENESIS
Most bacterial infections are acquired from an external source. However, some bacterial infections are caused by members of the normal flora and, as such, are not transmitted directly prior to the onset of infection.
A generalized sequence of the stages of infection is as follows:
(1) Transmission from an external source into the portal of entry.
(2) Evasion of primary host defenses such as skin or stomach acid.
(3) Adherence to mucous membranes, usually by bacterial pili.
(4) Colonization by growth of the bacteria at the site of adherence.
(5) Disease symptoms caused by toxin production or invasion accompanied by inflammation.
(6) Host responses, both nonspecific and specific (immunity), during steps 3, 4, and 5.
(7) Progression or resolution of the disease.
DETERMINANTS OF BACTERIAL PATHOGENESIS
1. Transmission
An understanding of the mode of transmission of bacteria and other infectious agents is extremely important from a public health perspective, because interrupting the chain of transmission is an excellent way to prevent infectious diseases. The mode of transmission of many infectious diseases is “human-to-human,” but infectious diseases are also transmitted from nonhuman sources such as soil, water, and animals. Fomites are inanimate objects, such as towels, that serve as a source of microorganisms that can cause infectious diseases. Table 7–1 describes some important examples of these modes of transmission.
TABLE 7–1 Important Modes of Transmission
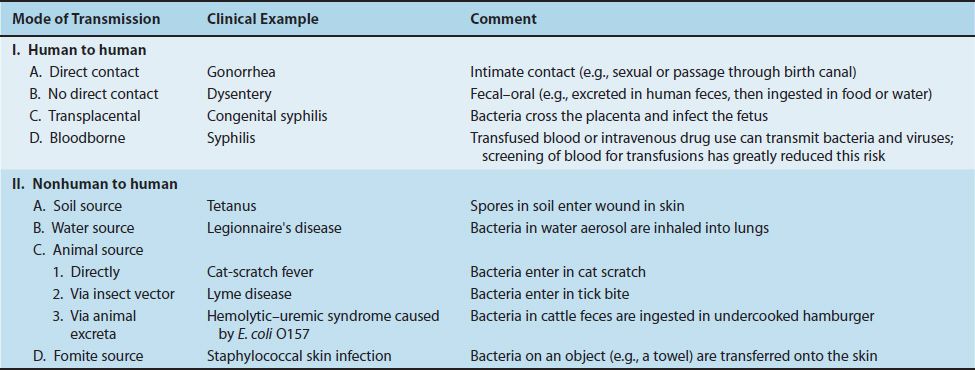
Although some infections are caused by members of the normal flora, most are acquired by transmission from external sources. Pathogens exit the infected patient most frequently from the respiratory and gastrointestinal tracts; hence transmission to the new host usually occurs via airborne respiratory droplets or fecal contamination of food and water. Organisms can also be transmitted by sexual contact, urine, skin contact, blood transfusions, contaminated needles, or biting insects. The transfer of blood, either by transfusion or by sharing needles during intravenous drug use, can transmit various bacterial and viral pathogens. The screening of donated blood for Treponema pallidum, human immunodeficiency virus (HIV), human T-cell lymphotropic virus, hepatitis B virus, hepatitis C virus, and West Nile virus has greatly reduced the risk of infection by these organisms.
The major bacterial diseases transmitted by ticks in the United States are Lyme disease, Rocky Mountain spotted fever, ehrlichiosis, relapsing fever, and tularemia. Of these five diseases, Lyme disease is by far the most common. Ticks of the genus Ixodes transmit three infectious diseases: Lyme disease, ehrlichiosis, and babesiosis, a protozoan disease.
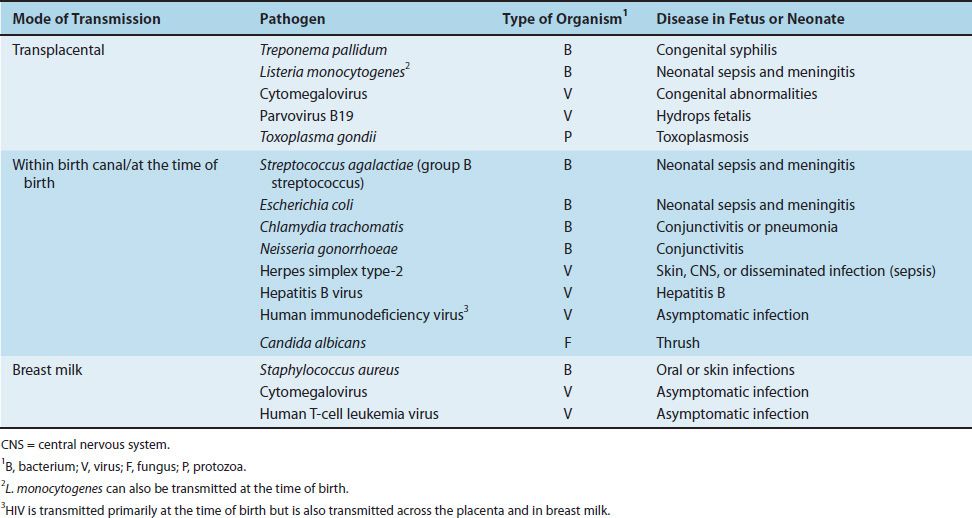
Bacteria, viruses, and other microbes can also be transmitted from mother to offspring, a process called vertical transmission. The three modes by which organisms are transmitted vertically are across the placenta, within the birth canal during birth, and via breast milk. Table 7–2 describes some medically important organisms that are transmitted vertically. (Horizontal transmission, by contrast, is person-to-person transmission that is not from mother to offspring.)
TABLE 7–2 Vertical Transmission of Some Important Pathogens
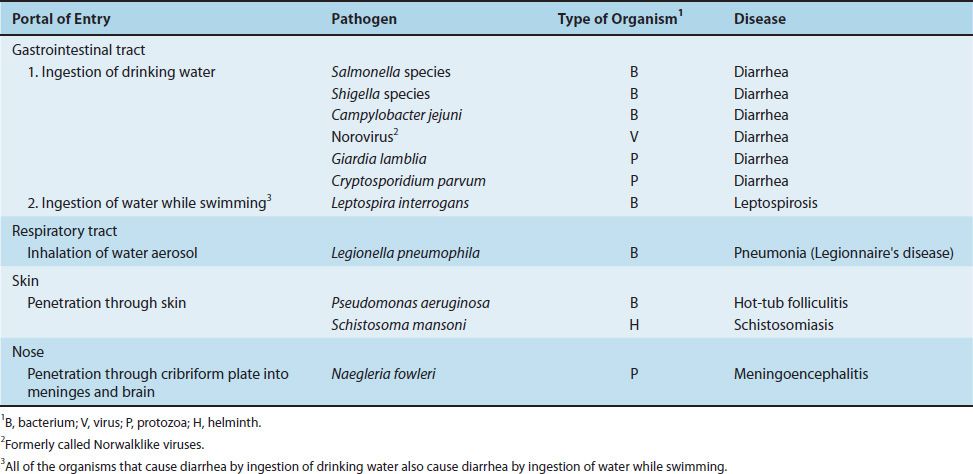
There are four important portals of entry: respiratory tract, gastrointestinal tract, genital tract, and skin (Table 7–3). Important microorganisms and diseases transmitted by water are described in Table 7–4.
TABLE 7–3 Portals of Entry of Some Common Pathogens
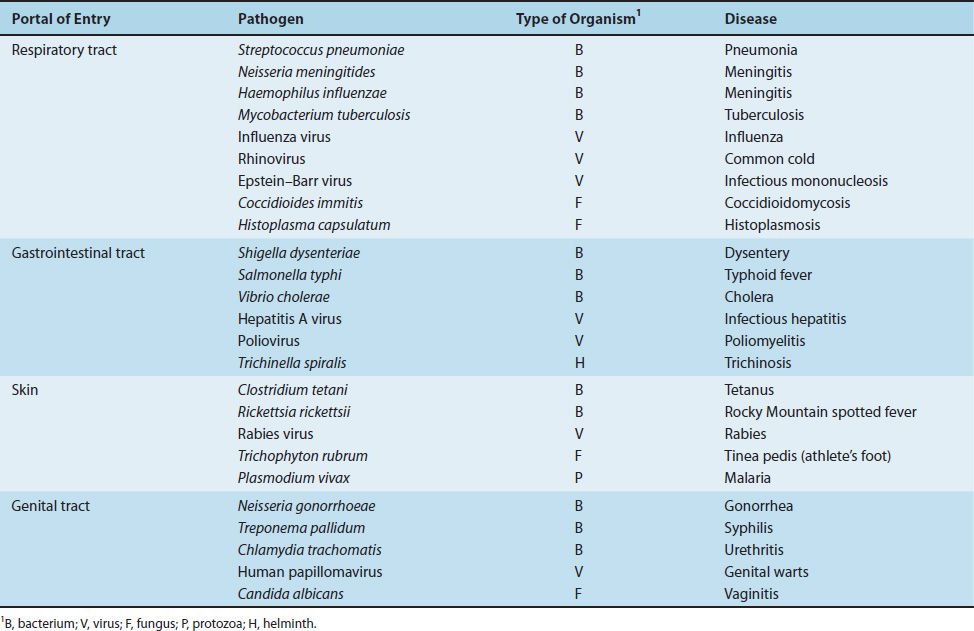
TABLE 7–4 Transmission of Important Waterborne Diseases
The important bacterial diseases transmitted by foods are listed in Table 7–5, and those transmitted by insects are listed in Table 7–6. The specific mode of transmission of each organism is described in the subsequent section devoted to that organism.
TABLE 7–5 Bacterial Diseases Transmitted by Foods
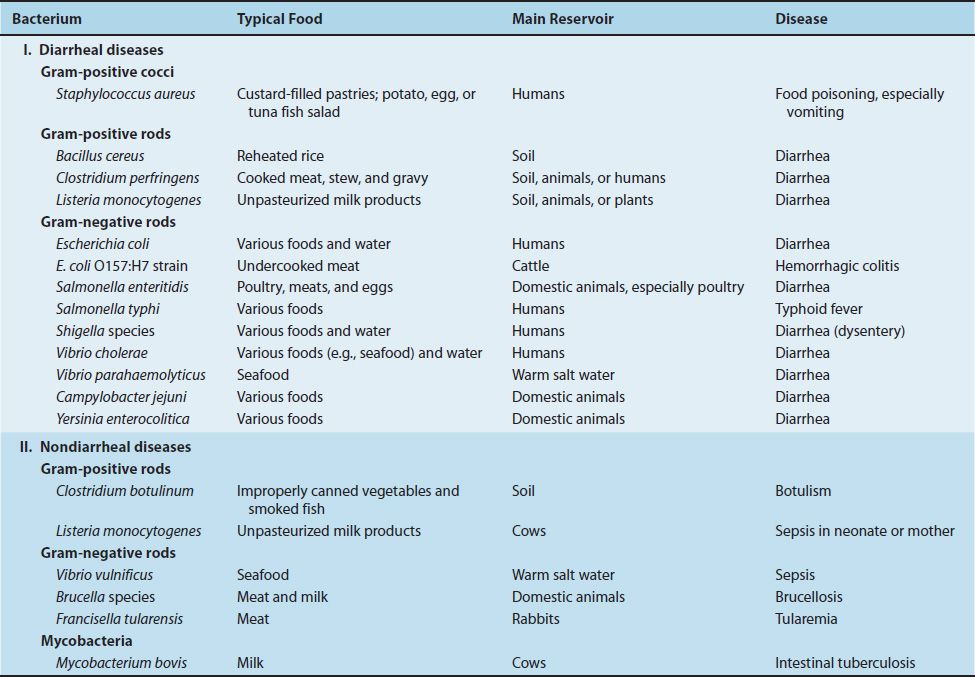
TABLE 7–6 Bacterial Diseases Transmitted by Insects
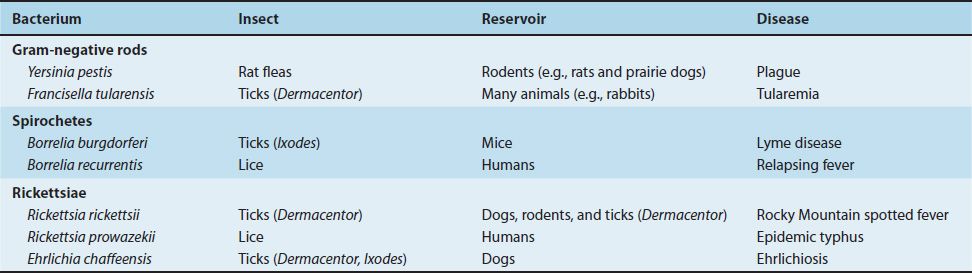
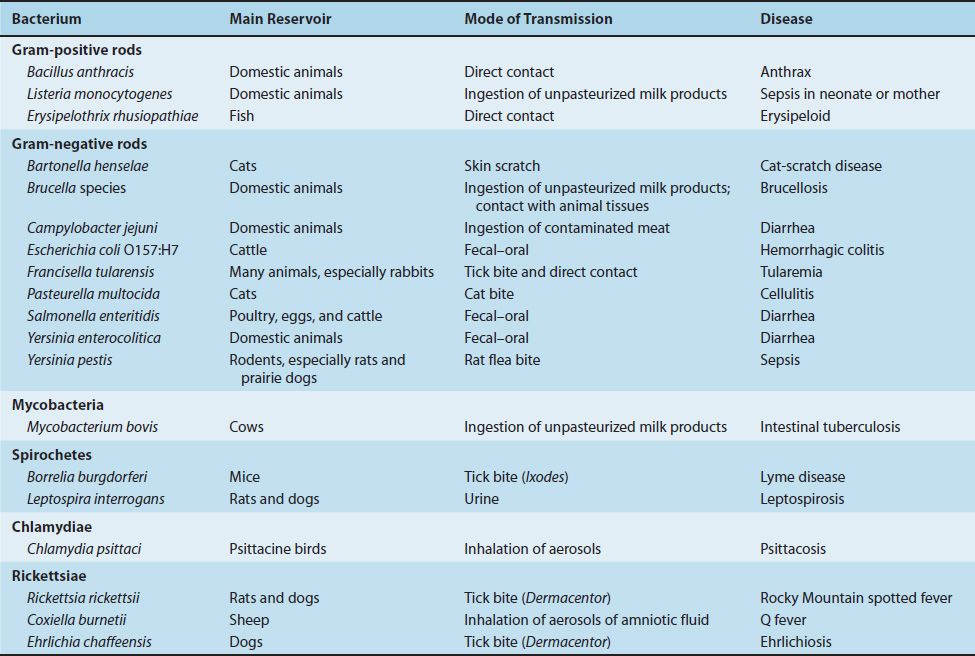
Animals are also an important source of organisms that infect humans. They can be either the source (reservoir) or the mode of transmission (vector) of certain organisms. Diseases for which animals are the reservoirs are called zoonoses. The important zoonotic diseases caused by bacteria are listed in Table 7–7.
TABLE 7–7 Zoonotic Diseases Caused by Bacteria
2. Adherence to Cell Surfaces
Certain bacteria have specialized structures (e.g., pili) or produce substances (e.g., capsules or glycocalyces) that allow them to adhere to the surface of human cells, thereby enhancing their ability to cause disease. These adherence mechanisms are essential for organisms that attach to mucous membranes; mutants that lack these mechanisms are often nonpathogenic. For example, the pili of Neisseria gonorrhoeae and E. coli mediate the attachment of the organisms to the urinary tract epithelium, and the glycocalyx of Staphylococcus epidermidis and certain viridans streptococci allows the organisms to adhere strongly to the endothelium of heart valves. The various molecules that mediate adherence to cell surfaces are called adhesins.
After the bacteria attach, they often form a protective matrix called a biofilm consisting of various polysaccharides and proteins. Biofilms form especially on foreign bodies such as prosthetic joints, prosthetic heart valves, and intravenous catheters, but they also form on native structures such as heart valves. Biofilms protect bacteria from both antibiotics and host immune defenses such as antibodies and neutrophils. They also retard wound healing resulting in chronic wound infections, especially in diabetics. Biofilms play an important role in the persistence of Pseudomonas in the lungs of cystic fibrosis patients and in the formation of dental plaque, the precursor of dental caries.
The production of biofilms by bacteria such as Pseudomonas is controlled by the process of quorum sensing. In quorum sensing, the bacteria grow in a nonaggressive manner until a quorum is sensed (i.e., a certain density of bacteria has been reached), at which point the synthesis of new bacterial virulence factors (e.g., biofilms) that contribute to pathogenesis occurs.
Foreign bodies, such as artificial heart valves and artificial joints, predispose to infections. Bacteria can adhere to these surfaces, but phagocytes adhere poorly owing to the absence of selectins and other binding proteins on the artificial surface (see Chapter 8).
Some strains of E. coli and Salmonella have surface proteins called curli, which mediate binding of the bacteria to endothelium and to extracellular proteins such as fibronectin. Curli also interact with serum proteins such as factor XII—a component of the coagulation cascade. Curli, therefore, are thought to play a role in the production of the thrombi seen in the disseminated intravascular coagulation (DIC) associated with sepsis caused by these bacteria. (See the discussion of endotoxin on page 44.)
3. Invasion, Inflammation, & Intracellular Survival
One of the two main mechanisms by which bacteria cause disease is invasion of tissue followed by inflammation. (The inflammatory response is described in Chapter 8.) The other main mechanism, toxin production, and a third mechanism, immunopathogenesis, are described later in this chapter.
Several enzymes secreted by invasive bacteria play a role in pathogenesis. Among the most prominent are the following:
(1) Collagenase and hyaluronidase, which degrade collagen and hyaluronic acid, respectively, thereby allowing the bacteria to spread through subcutaneous tissue; they are especially important in cellulitis caused by Streptococcus pyogenes.
(2) Coagulase, which is produced by Staphylococcus aureus and accelerates the formation of a fibrin clot from its precursor, fibrinogen (this clot may protect the bacteria from phagocytosis by walling off the infected area and by coating the organisms with a layer of fibrin). Coagulase is also produced by Yersinia pestis, the cause of bubonic plague. See Chapter 20 for the role of coagulase in the pathogenesis of plague.
(3) Immunoglobulin A (IgA) protease, which degrades IgA, allowing the organism to adhere to mucous membranes, and is produced chiefly by N. gonorrhoeae, Haemophilus influenzae, and Streptococcus pneumoniae.
(4) Leukocidins, which can destroy both neutrophilic leukocytes and macrophages.
In addition to these enzymes, several virulence factors contribute to invasiveness by limiting the ability of the host defense mechanisms, especially phagocytosis, to operate effectively.
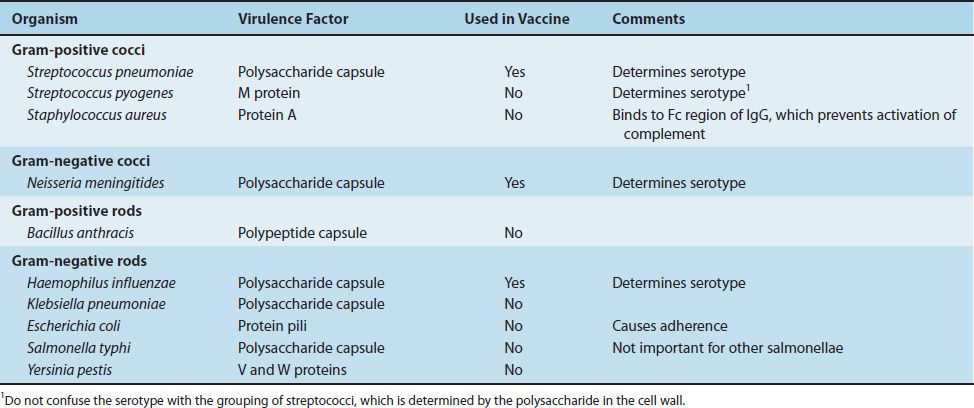
(1) The most important of these antiphagocytic factors is the capsule external to the cell wall of several important pathogens such as Str. pneumoniae and Neisseria meningitidis. The polysaccharide capsule prevents the phagocyte from adhering to the bacteria; anticapsular antibodies allow more effective phagocytosis to occur (a process called opsonization) (see page 55). The vaccines against Str. pneumoniae, H. influenzae, and N. meningitidis contain capsular polysaccharides that induce protective anticapsular antibodies.
(2) A second group of antiphagocytic factors are the cell wall proteins of the gram-positive cocci, such as the M protein of the group A streptococci (Str. pyogenes) and protein A of Sta. aureus. The M protein is antiphagocytic, and protein A binds to immunoglobulin G (IgG) and prevents the activation of complement. These virulence factors are summarized in Table 7–8.
TABLE 7–8 Surface Virulence Factors Important for Bacterial Pathogenesis
Bacteria can cause two types of inflammation: pyogenic and granulomatous. In pyogenic (pus-producing) inflammation, neutrophils are the predominant cells. Some of the most important pyogenic bacteria are the gram-positive and gram-negative cocci listed in Table 7–8. In granulomatous inflammation, macrophages and T cells predominate. The most important organism in this category is Mycobacterium tuberculosis. No bacterial enzymes or toxins that induce granulomas have been identified. Rather, it appears that bacterial antigens stimulate the cell-mediated immune system, resulting in sensitized T-lymphocyte and macrophage activity. Phagocytosis by macrophages kills most of the bacteria, but some survive and grow within the macrophages in the granuloma.
Intracellular survival is an important attribute of certain bacteria that enhances their ability to cause disease. These bacteria are called “intracellular” pathogens and commonly cause granulomatous lesions. The best-known of these bacteria belong to the genera Mycobacterium, Legionella, Brucella, and Listeria. The best-known fungus is Histoplasma. These organisms are not obligate intracellular parasites, which distinguishes them from Chlamydia and Rickettsia. They can be cultured on microbiologic media in the laboratory and therefore are not obligate intracellular parasites. Rather, they prefer an intracellular location probably because they are protected there from antibody and neutrophils that function extracellularly.
These bacteria use several different mechanisms to allow them to survive and grow intracellularly. These include (1) inhibition of the fusion of the phagosome with the lysosome, which allows the organisms to avoid the degradative enzymes in the lysosome; (2) inhibition of acidification of the phagosome, which reduces the activity of the lysosomal degradative enzymes; and (3) escape from the phagosome into the cytoplasm, where there are no degradative enzymes. Members of the genera Mycobacterium and Legionella are known to use the first and second mechanisms, whereas Listeria species use the third.
The invasion of cells by bacteria is dependent on the interaction of specific bacterial surface proteins called invasins and specific cellular receptors belonging to the integrin family of transmembrane adhesion proteins. The movement of bacteria into the cell is a function of actin microfilaments. Once inside the cell, these bacteria typically reside within cell vacuoles such as phagosomes. Some remain there, others migrate into the cytoplasm, and some move from the cytoplasm into adjacent cells through tunnels formed from actin. Infection of the surrounding cells in this manner allows the bacteria to evade host defenses. For example, Listeria monocytogenes aggregates actin filaments on its surface and is propelled in a “sling-shot” fashion, called actin rockets, from one host cell to another.
The “Yops” (Yersinia outer-membrane proteins) produced by several Yersinia species are important examples of bacterial virulence factors that act primarily after invasion of human cells by the organism. The most important effects of the Yops proteins are to inhibit phagocytosis by neutrophils and macrophages and to inhibit cytokine production (e.g., tumor necrosis factor [TNF] production) by macrophages. For example, one of the Yops proteins of Yersinia pestis (Yop J) is a protease that cleaves signal transduction proteins required for the induction of TNF synthesis. This inhibits the activation of our host defenses and contributes to the ability of the organism to cause bubonic plague.
The genes that encode many virulence factors in bacteria are clustered in pathogenicity islands on the bacterial chromosome. For example, in many bacteria, the genes encoding adhesins, invasins, and exotoxins are adjacent to each other on these islands. Nonpathogenic variants of these bacteria do not have these pathogenicity islands. It appears that these large regions of the bacterial genome were transferred as a block via conjugation or transduction. Unlike plasmids and bacteriophage, pathogenicity islands do not have the ability to replicate independent of the bacterial chromosome. Pathogenicity islands are found in many gram-negative rods, such as E. coli, Salmonella, Shigella, Pseudomonas, and Vibrio cholerae, and in gram-positive cocci, such as S. pneumoniae. Additional information about pathogenicity islands is given on page 47.
After bacteria have colonized and multiplied at the portal of entry, they may invade the bloodstream and spread to other parts of the body. Receptors for the bacteria on the surface of cells determine, in large part, the organs affected. For example, certain bacteria or viruses infect the brain because receptors for these microbes are located on the surface of brain neurons. The blood–brain barrier, which limits the ability of certain drugs to penetrate the brain, is not thought to be a determinant of microbial infection of the brain. The concept of a blood–brain barrier primarily refers to the inability of hydrophilic (charged, ionized) drugs to enter the lipid-rich brain parenchyma, whereas lipophilic (lipid-soluble) drugs enter well.
Two important diseases, diphtheria and pseudomembranous colitis, are characterized by inflammatory lesions called pseudomembranes. Pseudomembranes are thick, adherent, grayish or yellowish exudates on the mucosal surfaces of the throat in diphtheria and on the colon in pseudomembranous colitis. The term pseudo refers to the abnormal nature of these membranes in contrast to the normal anatomic membranes of the body, such as the tympanic membrane and the placental membranes.
4. Toxin Production
The second major mechanism by which bacteria cause disease is the production of toxins. A comparison of the main features of exotoxins and endotoxins is shown in Table 7–9.
TABLE 7–9 Main Features of Exotoxins and Endotoxins
Exotoxins
Exotoxins are produced by several gram-positive and gram-negative bacteria, in contrast to endotoxins, which are present only in gram-negative bacteria. The essential characteristic of exotoxins is that they are secreted by the bacteria, whereas endotoxin is a component of the cell wall. Exotoxins are polypeptides whose genes are frequently located on plasmids or lysogenic bacterial viruses (bacteriophages). Some important exotoxins encoded by bacteriophage DNA are diphtheria toxin, cholera toxin, and botulinum toxin.
Exotoxins are among the most toxic substances known. For example, the fatal dose of tetanus toxin for a human is estimated to be less than 1 μg. Because some purified exotoxins can reproduce all aspects of the disease, we can conclude that certain bacteria play no other role in pathogenesis than to synthesize the exotoxin. Exotoxin polypeptides are good antigens and induce the synthesis of protective antibodies called antitoxins, some of which are useful in the prevention or treatment of diseases such as botulism and tetanus. When treated with formaldehyde (or acid or heat), the exotoxin polypeptides are converted into toxoids, which are used in protective vaccines because they retain their antigenicity but have lost their toxicity.
Many exotoxins have an A–B subunit structure; the A (or active) subunit possesses the toxic activity, and the B (or binding) subunit is responsible for binding the exotoxin to specific receptors on the membrane of the human cell. The binding of the B subunit determines the specific site of the action of the exotoxin. For example, botulinum toxin acts at the neuromuscular junction because the B subunit binds to specific receptors on the surface of the motor neuron at the junction. Important exotoxins that have an A–B subunit structure include diphtheria toxin, tetanus toxin, botulinum toxin, cholera toxin, and the enterotoxin of E. coli (Figure 7–1).
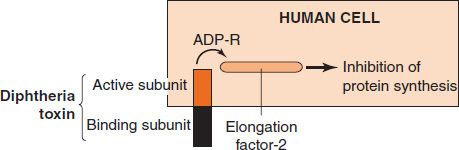
FIGURE 7–1 Mode of action of diphtheria toxin. The toxin binds to the cell surface via its binding subunit, and the active subunit enters the cell. The active subunit is an enzyme that catalyzes the addition of ADP-ribose (ADP-R) to elongation factor-2 (EF-2). This inactivates EF-2, and protein synthesis is inhibited.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


