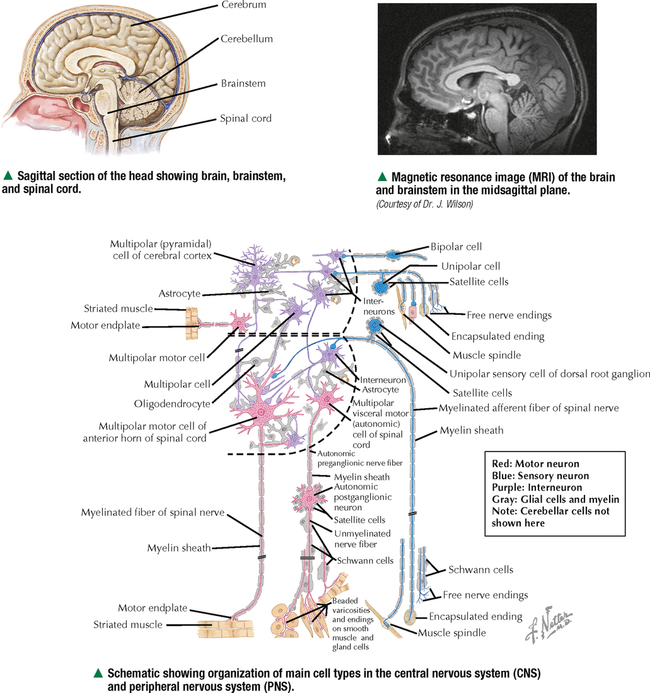
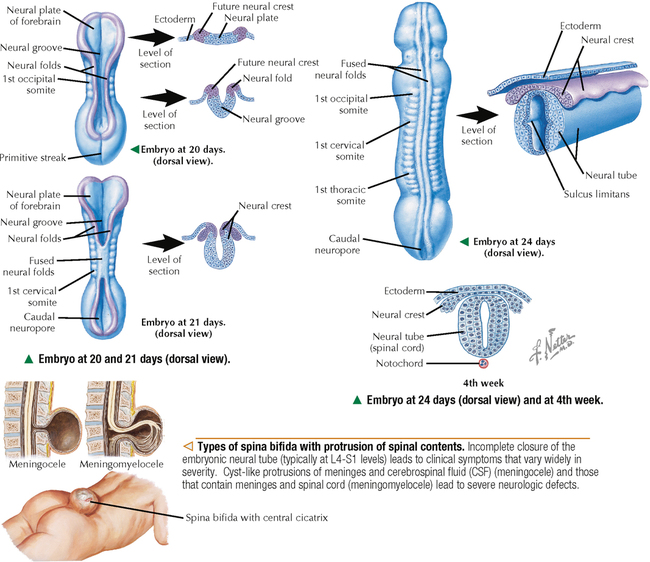
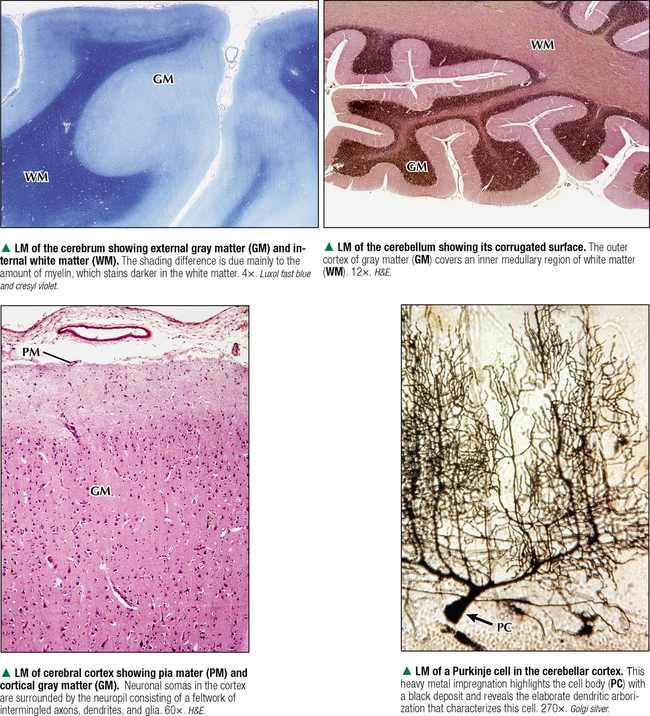
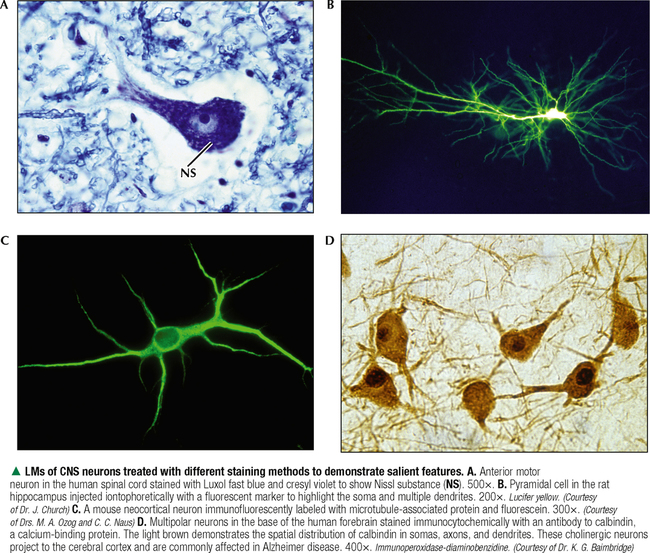
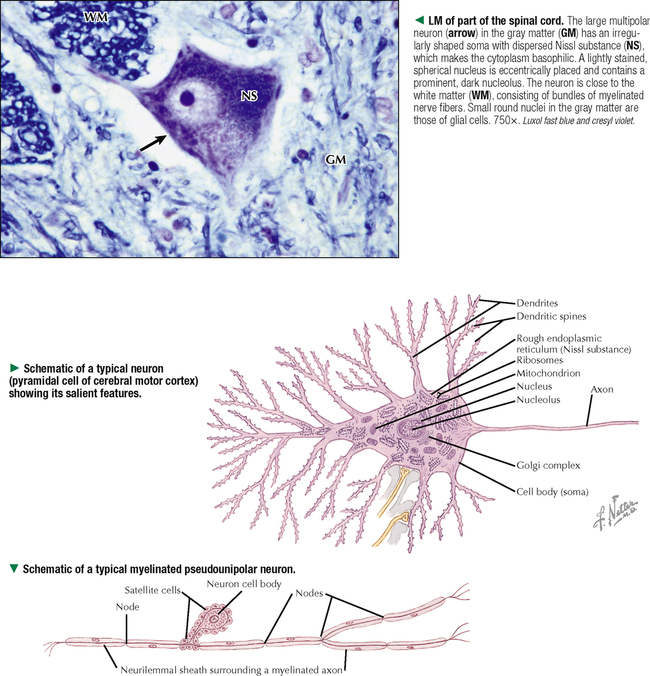
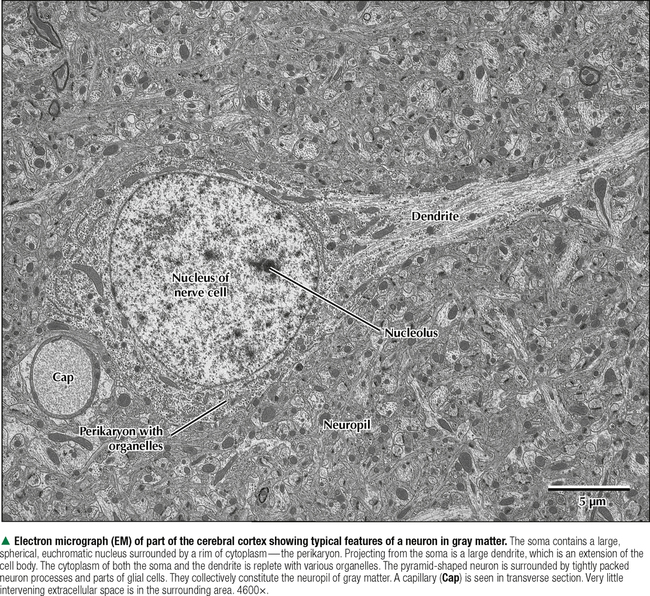
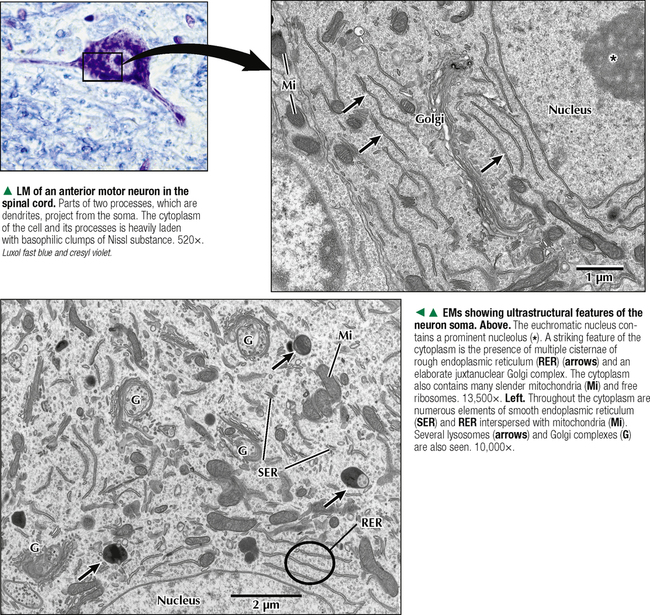
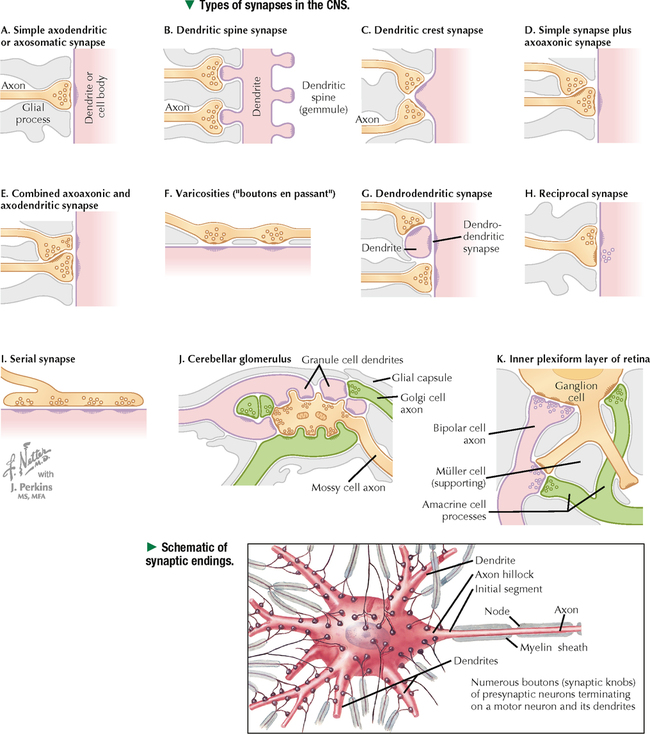
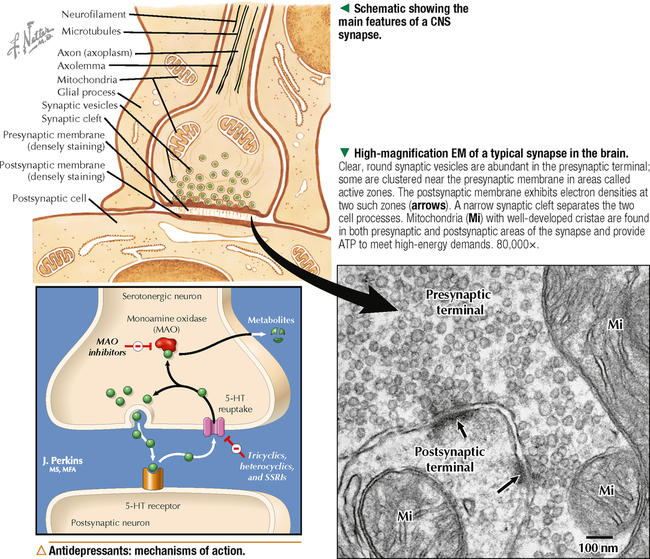
5 NERVOUS TISSUE 5.1. Overview 5.2. Embryonic Development 5.3. Structure and Function of the Meninges 5.4. Neurocytology: Cytoarchitecture 5.5. Neurocytology: Staining Methods 5.6. Structure of a Neuron 5.7. Ultrastructure of a Neuron in Gray Matter in Relation to Surrounding Structures 5.8. Ultrastructure of a Spinal Cord Neuron Soma 5.9. Types of Synapses 5.10. Ultrastructure of Synapses 5.11. Structure and Function of Glial Cells 5.12. Structure and Function of Astrocytes 5.13. Structure and Function of the Blood-Brain Barrier 5.14. Ultrastructure of the Blood-Brain Barrier 5.15. Myelination of Axons in the Central and Peripheral Nervous Systems 5.16. Oligodendrocytes and Myelination in the Central Nervous System 5.17. Structure and Function of the Ependyma 5.18. Structure and Function of the Choroid Plexus 5.19. Cytoarchitecture of the Cerebral Cortex 5.20. Cytoarchitecture of the Cerebellum 5.21. Histology and Ultrastructure of the Cerebellum 5.22. Anatomy and Histology of the Spinal Cord 5.23. Histology of Peripheral Nerves 5.24. Ultrastructure of Myelinated and Unmyelinated Nerve Fibers in the Peripheral Nervous System 5.25. Ultrastructure of Myelinated Nerve Fibers in the Peripheral Nervous System 5.26. Nerve Fibers in Longitudinal Section and Nodes of Ranvier in the Peripheral Nervous System 5.27. Histology of Peripheral Autonomic Ganglia 5.28. Histology and Ultrastructure of Peripheral Ganglia 5.29. Ultrastructure and Function of the Perineurium 5.1 OVERVIEW The nervous system is divided anatomically into the central (CNS) and peripheral (PNS) nervous systems. The CNS comprises the brain, brainstem, and spinal cord; the PNS, all nerve fibers (axons and dendrites), nerve endings, and collections of their cell bodies that lie outside the CNS. The autonomic nervous system, a subdivision of the PNS, is connected to the CNS through spinal and cranial nerves. Its sympathetic and parasympathetic portions innervate organs and tissues that are under autonomic, or involuntary, control such as glands, smooth muscle, and cardiac muscle. The CNS and PNS contain nervous tissue, one of the four basic body tissues, which possesses two major cell types: nerve cells, or neurons, and supporting cells, or glia. Neurons can generate nervous impulses in response to stimuli and transmit them along cellular processes. More than 50 billion neurons are estimated to be in the nervous system. The types of neurons are classified on the basis of appearance, shape, and number of processes as multipolar, bipolar, or pseudounipolar. Despite their variability, all neurons conform to a common histologic plan: highly specialized cells with several parts to carry out functions of receiving signals and then transmitting information as nerve impulses to other neurons or effector organs. Conductivity and irritability are best developed in neurons; glial cells are non-impulseconducting cells that represent interstitial tissue and mostly support and protect neurons. 5.2 EMBRYONIC DEVELOPMENT The nervous system develops from a thickening of dorsal ectoderm of the early embryo. At 14–16 days, a neural plate of surface ectoderm appears in the dorsal midline. It becomes indented and forms a longitudinal neural groove with neural folds on each side. By 24 days, the neural folds fuse dorsally to form a neural tube, which becomes the CNS, including the brain rostrally and spinal cord more caudally. The neural tube is first open at both ends, but at 24–26 days it closes. Isolated cells not incorporated into the neural tube form a strip of neuroectodermal cells—the neural crest. These cells migrate ventrolaterally along each side of the neural tube to form a series of somites. The neural crest ectoderm ultimately gives rise to PNS components, including dorsal root ganglia of spinal nerves, comparable sensory ganglia of cranial nerves, autonomic ganglia, and chromaffin cells of the adrenal medulla. Neuron bodies inside the CNS are derived from the neural tube; those outside, in the PNS, the neural crest. Axons and dendrites sprout from neuron bodies and grow long distances. Supporting satellite cells envelop neuron bodies in the PNS, whereas cells around neuron peripheral processes or nerve fibers are Schwann cells. The neural tube lumen gives rise to fluid-filled ventricles of the brain and central canal of the spinal cord. Coverings of the brain and spinal cord, known as meninges, develop later. They are composed of three distinct layers: an outermost dura mater, arachnoid, and innermost pia mater. CLINICAL POINT Malformations of the developing nervous system may arise during closure and later growth of the neural tube and result in various neural tube defects. Anencephaly is a congenital malformation caused by failure of fusion of neural folds in rostral regions. Degeneration of unfused folds leads to failure of development of neural tissue and absence of most of the brain, the result being stillbirth or premature death. A defect at more caudal levels of the primitive spinal cord is called spina bifida. This condition typically produces paralysis depending on the level of the lesion and is usually not life-threatening. 5.3 STRUCTURE AND FUNCTION OF THE MENINGES The term meninges derives from the Greek meninx, meaning membrane. The three layers of the meninges stabilize and protect the brain and spinal cord. The dura mater invests the brain, spinal cord, and optic nerves. The thickest and toughest layer, the dura, is dense, fibrous connective tissue consisting of interlacing bundles of collagen and elastic fibers associated with flattened fibroblasts. The outer aspect of the dura attaches to the periosteum of the skull; the inner dural surface is lined by a layer of flattened fibroblasts. The dura contains large blood vessels, nerves, and lymphatics. Two potential spaces associated with it are the epidural space (exterior) and subdural space (between dura and arachnoid). These normally potential spaces can in some pathologic conditions accumulate fluid such as blood. The arachnoid and pia mater are thinner and more delicate than the dura and are known together as the leptomeninges. The arachnoid comprises several layers of flattened, closely packed fibroblasts linked by tight junctions with some intervening collagen. The arachnoid, so named because it resembles a spider’s web, sends inward projections, the arachnoid trabeculae, into the subarachnoid space to form a “cobweb” that merges with the pia. This space is filled with cerebrospinal fluid (CSF) and contains branches of cerebral arteries and veins. Peripherally, the arachnoid is continuous with the perineurium around peripheral nerve fascicles. The pia mater intimately invests all external surfaces of the CNS and extends into its folds, fissures, and convolutions. At certain sites, the pia protrudes into the ventricles close to modified ependymal cells to form the choroid plexus. CLINICAL POINT Meningitis, or inflammation of the meninges, is most often caused by bacteria or viruses. Other pathogens such as fungi or parasites are also causes. Bacterial meningitis is less common than the viral form, can be life-threatening, and is characterized by exudates of polymorphonuclear leukocytes in the CNS. Viral hepatitis is marked mostly by lymphocyte infiltration in the brain and raised numbers of T cells in CSF. A leading cause of meningitis in children is Haemophilus influenzae type b, and a vaccine for it has dramatically reduced its incidence. Meningitis may occur at any age, but it is most common in children, the elderly, and immunocompromised people. 5.4 NEUROCYTOLOGY: CYTOARCHITECTURE The unique cytoarchitecture of the CNS varies regionally. By inspection with the naked eye, the CNS is made up of gray matter and white matter. Unmyelinated neuronal processes and glial cells, tracts of myelinated nerve fibers, and associated glia dominate the white matter, whereas gray matter consists mostly of neuron bodies. The CNS also has a rich vascular supply that includes a profuse network of capillaries, which are more abundant in gray matter. In the spinal cord, gray matter is located internally and is enveloped by an external layer of white matter. In other regions of the CNS, such as cerebrum and cerebellum, an outer cortex of gray matter covers an internal medullary region of white matter. Because of the complexity and intricate nature of nervous tissue, ordinary staining methods have limited value when used alone to examine its cytologic features. 5.5 NEUROCYTOLOGY: STAINING METHODS Special techniques with varied and selective stains, which provide a composite view of nervous tissue, are often used in neurohistology and neuropathology. Basic cationic dyes such as cresyl violet and toluidine blue elucidate cell nuclei and neuronal Nissl substance (rough endoplasmic reticulum and ribosomes). Luxol fast blue and other reagents such as osmium can demonstrate myelin sheaths. Metal impregnation with reduced gold and silver demonstrates the intricate nature of axons and dendrites. Histochemistry and immunocytochemistry permit localization of specific substances and molecules within different types of neurons and glia. Electron microscopy has proved to be quite useful for revealing fine structural details beyond the resolution power of light microscopy. CLINICAL POINT Senile dementia of the Alzheimer type, or Alzheimer disease, which is characterized by progressive memory loss, is increasingly common in developed countries as populations include more elderly persons. Definitive diagnosis is made by microscopic examination of the brain at autopsy, with the histopathologic hallmark being an increased number of neuritic, or senile, plaques, which occur mostly in the cerebral cortex. They consist of tortuous neuritic processes formed mainly by degenerative presynaptic endings, which surround a central amyloid core. Reactive astrocytes and microglia may appear at a plaque periphery. Another important histologic feature is proliferation of intracytoplasmic neurofibrillary tangles. 5.6 STRUCTURE OF A NEURON The neuron is a highly polarized cell that consists of a soma, or cell body, from which cytoplasmic processes arise. The processes, known as nerve fibers, vary greatly in size, some being up to 1.5 m long. Processes conducting impulses toward cell bodies are dendrites, whereas a single process conveying impulses away from cell bodies is an axon. The soma consists of a nucleus and the surrounding cytoplasm, known as the perikaryon. Soma sizes, which depend on cell type and function, vary from 5 to 150 μm. Anterior motor neurons of the spinal cord are among the largest in the CNS, whereas granule cells in the cerebellar cortex are among the smallest. Irregularly shaped masses of basophilic material known as Nissl substance are scattered in the cytoplasm of the body and the dendrites. Neurons are classified into three types on the basis of the number of processes. Multipolar neurons are the most common and characteristic and have one axon and several dendrites. Bipolar neurons have two processes, an axon and a dendrite, and are found in the visual, auditory, and olfactory systems. Pseudounipolar neurons have one short process, which bifurcates into an axon and a dendrite. 5.7 ULTRASTRUCTURE OF A NEURON IN GRAY MATTER IN RELATION TO SURROUNDING STRUCTURES The soma is the trophic center of the neuron and varies greatly in size and shape. Reflecting its role in genetic regulation and transcription, the euchromatic nucleus of a typical neuron has small patches of heterochromatin, peripherally displaced, just under the nuclear envelope. The nucleus is usually spherical to ovoid, and it is large relative to the surrounding perikaryon. A characteristic feature is one or more prominent nucleoli, seen in fortuitous sections; they have a role in synthesizing ribosomal RNA. The surrounding cytoplasm is the site of synthesis of most of the structural and secretory proteins, enzymes, and organelles needed for diverse functions of the cell. Dominating the cytoplasm are multiple flattened cisternae of rough endoplasmic reticulum between which are numerous free ribosomes (the basophilic Nissl substance seen in light micrographs). Their major function is protein synthesis for internal use and export. A highly developed Golgi complex with multiple stacks of flattened sacs and associated vesicles and vacuoles is usually seen near the nucleus. This organelle is responsible for packaging and concentration of secretory products, posttranslational modification of macromolecules, and supply of lysosomes, which accumulate wear-and-tear lipofuscin pigment in these long-lived cells with advancing age. 5.8 ULTRASTRUCTURE OF A SPINAL CORD NEURON SOMA Mitochondria, the source of ATP to meet energy requirements for the metabolically active neuron, are abundant throughout the soma and in peripheral processes of the cell. They are especially numerous in axon terminals close to synapses. Neuronal cytoplasm has a well-developed cytoskeleton consisting of microtubules, actin filaments, and neurofilaments (an intermediate type). These organelles, found throughout the soma and extending into the axon and dendrites, help maintain cell shape and structural stability. Microtubules provide intracellular axoplasmic transport of organelles, most notably of mitochondria and membrane-bound vesicles containing precursors of neurotransmitters. Intracellular neuronal transport is bidirectional: anterograde transport is directed away from the soma and into cytoplasmic processes, whereas retrograde transport transports organelles and other material toward the soma. Microtubules and neurofilaments also play roles in axonal growth and guidance during development and in regeneration after injury. Neurofilaments are best seen by electron microscopy and can be detected by immunocytochemistry, but an affinity for heavy metals is the basis for metal impregnation staining techniques developed more than a century ago. Neuronal cytoplasm also contains lipid droplets, smooth endoplasmic reticulum, lysosomes, peroxisomes, Golgi complexes, ribosomes, and sometimes melanin pigment granules. 5.9 TYPES OF SYNAPSES Synapses are specialized sites for chemical or electrical transmission for communication between neurons or between neurons and other effector cells such as skeletal muscle fibers. Most synapses in humans involve chemical neurotransmitters, which are released from presynaptic terminals of one axon or dendrite to affect receptors on the postsynaptic membrane of the target cell. Various neurotransmitters exist and include amino acids such as glutamate, catecholamines such as epinephrine and norepinephrine, serotonin, neuropeptides, and acetylcholine. In functional terms, two main types of synapses occur: excitatory and inhibitory. In excitatory synapses, neurotransmitter release from the presynaptic neuron depolarizes the postsynaptic membrane; in inhibitory synapses, the postsynaptic membrane is hyperpolarized. Most CNS synapses are between an axon of one neuron and the dendrite of another—axodendritic. Other types include axosomatic and, less commonly, axoaxonic synapses. In some sites, such as hypothalamus and posterior pituitary, large vesicles in presynaptic terminals may contain polypeptide hormones, for example, oxytocin or vasopressin, that are neurosecretory products, not neurotransmitters. 5.10 ULTRASTRUCTURE OF SYNAPSES A typical synapse in the CNS consists of three major components: presynaptic terminal, synaptic cleft, and postsynaptic membrane. The presynaptic terminal aligns closely with the postsynaptic membrane of the target cell. In the area of membrane apposition, presynaptic and postsynaptic membranes are separated by a narrow synaptic cleft 12–30 nm wide. Clusters of large numbers of synaptic vesicles in the presynaptic terminal contain neurotransmitter that is released by exocytosis to mediate synaptic transmission. By electron microscopy, synaptic vesicles are 40–60 nm in diameter and are membrane-bound. Whether they have a clear center or an electron-dense core depends on the chemical nature of the neurotransmitter. Pre- and postsynaptic membrane specializations contain electron-dense material that extends into underlying cytoplasm and is usually thicker in the postsynaptic area. An action potential causes presynaptic vesicles to fuse with the presynaptic membrane and discharge neurotransmitter into the synaptic cleft. Neurotransmitter then diffuses across the cleft to interact with receptor molecules on the postsynaptic membrane, which changes postsynaptic membrane conductance Only gold members can continue reading. Log In or Register to continue Share this: Share on X (Opens in new window) X Share on Facebook (Opens in new window) Facebook Like this:Like Loading… Related Related posts: CARDIOVASCULAR SYSTEM RESPIRATORY SYSTEM SPECIAL SENSES FEMALE REPRODUCTIVE SYSTEM Stay updated, free articles. Join our Telegram channel Join Tags: Netters Essential Histology Jun 18, 2016 | Posted by admin in HISTOLOGY | Comments Off on NERVOUS TISSUE Full access? Get Clinical Tree
5 NERVOUS TISSUE 5.1. Overview 5.2. Embryonic Development 5.3. Structure and Function of the Meninges 5.4. Neurocytology: Cytoarchitecture 5.5. Neurocytology: Staining Methods 5.6. Structure of a Neuron 5.7. Ultrastructure of a Neuron in Gray Matter in Relation to Surrounding Structures 5.8. Ultrastructure of a Spinal Cord Neuron Soma 5.9. Types of Synapses 5.10. Ultrastructure of Synapses 5.11. Structure and Function of Glial Cells 5.12. Structure and Function of Astrocytes 5.13. Structure and Function of the Blood-Brain Barrier 5.14. Ultrastructure of the Blood-Brain Barrier 5.15. Myelination of Axons in the Central and Peripheral Nervous Systems 5.16. Oligodendrocytes and Myelination in the Central Nervous System 5.17. Structure and Function of the Ependyma 5.18. Structure and Function of the Choroid Plexus 5.19. Cytoarchitecture of the Cerebral Cortex 5.20. Cytoarchitecture of the Cerebellum 5.21. Histology and Ultrastructure of the Cerebellum 5.22. Anatomy and Histology of the Spinal Cord 5.23. Histology of Peripheral Nerves 5.24. Ultrastructure of Myelinated and Unmyelinated Nerve Fibers in the Peripheral Nervous System 5.25. Ultrastructure of Myelinated Nerve Fibers in the Peripheral Nervous System 5.26. Nerve Fibers in Longitudinal Section and Nodes of Ranvier in the Peripheral Nervous System 5.27. Histology of Peripheral Autonomic Ganglia 5.28. Histology and Ultrastructure of Peripheral Ganglia 5.29. Ultrastructure and Function of the Perineurium 5.1 OVERVIEW The nervous system is divided anatomically into the central (CNS) and peripheral (PNS) nervous systems. The CNS comprises the brain, brainstem, and spinal cord; the PNS, all nerve fibers (axons and dendrites), nerve endings, and collections of their cell bodies that lie outside the CNS. The autonomic nervous system, a subdivision of the PNS, is connected to the CNS through spinal and cranial nerves. Its sympathetic and parasympathetic portions innervate organs and tissues that are under autonomic, or involuntary, control such as glands, smooth muscle, and cardiac muscle. The CNS and PNS contain nervous tissue, one of the four basic body tissues, which possesses two major cell types: nerve cells, or neurons, and supporting cells, or glia. Neurons can generate nervous impulses in response to stimuli and transmit them along cellular processes. More than 50 billion neurons are estimated to be in the nervous system. The types of neurons are classified on the basis of appearance, shape, and number of processes as multipolar, bipolar, or pseudounipolar. Despite their variability, all neurons conform to a common histologic plan: highly specialized cells with several parts to carry out functions of receiving signals and then transmitting information as nerve impulses to other neurons or effector organs. Conductivity and irritability are best developed in neurons; glial cells are non-impulseconducting cells that represent interstitial tissue and mostly support and protect neurons. 5.2 EMBRYONIC DEVELOPMENT The nervous system develops from a thickening of dorsal ectoderm of the early embryo. At 14–16 days, a neural plate of surface ectoderm appears in the dorsal midline. It becomes indented and forms a longitudinal neural groove with neural folds on each side. By 24 days, the neural folds fuse dorsally to form a neural tube, which becomes the CNS, including the brain rostrally and spinal cord more caudally. The neural tube is first open at both ends, but at 24–26 days it closes. Isolated cells not incorporated into the neural tube form a strip of neuroectodermal cells—the neural crest. These cells migrate ventrolaterally along each side of the neural tube to form a series of somites. The neural crest ectoderm ultimately gives rise to PNS components, including dorsal root ganglia of spinal nerves, comparable sensory ganglia of cranial nerves, autonomic ganglia, and chromaffin cells of the adrenal medulla. Neuron bodies inside the CNS are derived from the neural tube; those outside, in the PNS, the neural crest. Axons and dendrites sprout from neuron bodies and grow long distances. Supporting satellite cells envelop neuron bodies in the PNS, whereas cells around neuron peripheral processes or nerve fibers are Schwann cells. The neural tube lumen gives rise to fluid-filled ventricles of the brain and central canal of the spinal cord. Coverings of the brain and spinal cord, known as meninges, develop later. They are composed of three distinct layers: an outermost dura mater, arachnoid, and innermost pia mater. CLINICAL POINT Malformations of the developing nervous system may arise during closure and later growth of the neural tube and result in various neural tube defects. Anencephaly is a congenital malformation caused by failure of fusion of neural folds in rostral regions. Degeneration of unfused folds leads to failure of development of neural tissue and absence of most of the brain, the result being stillbirth or premature death. A defect at more caudal levels of the primitive spinal cord is called spina bifida. This condition typically produces paralysis depending on the level of the lesion and is usually not life-threatening. 5.3 STRUCTURE AND FUNCTION OF THE MENINGES The term meninges derives from the Greek meninx, meaning membrane. The three layers of the meninges stabilize and protect the brain and spinal cord. The dura mater invests the brain, spinal cord, and optic nerves. The thickest and toughest layer, the dura, is dense, fibrous connective tissue consisting of interlacing bundles of collagen and elastic fibers associated with flattened fibroblasts. The outer aspect of the dura attaches to the periosteum of the skull; the inner dural surface is lined by a layer of flattened fibroblasts. The dura contains large blood vessels, nerves, and lymphatics. Two potential spaces associated with it are the epidural space (exterior) and subdural space (between dura and arachnoid). These normally potential spaces can in some pathologic conditions accumulate fluid such as blood. The arachnoid and pia mater are thinner and more delicate than the dura and are known together as the leptomeninges. The arachnoid comprises several layers of flattened, closely packed fibroblasts linked by tight junctions with some intervening collagen. The arachnoid, so named because it resembles a spider’s web, sends inward projections, the arachnoid trabeculae, into the subarachnoid space to form a “cobweb” that merges with the pia. This space is filled with cerebrospinal fluid (CSF) and contains branches of cerebral arteries and veins. Peripherally, the arachnoid is continuous with the perineurium around peripheral nerve fascicles. The pia mater intimately invests all external surfaces of the CNS and extends into its folds, fissures, and convolutions. At certain sites, the pia protrudes into the ventricles close to modified ependymal cells to form the choroid plexus. CLINICAL POINT Meningitis, or inflammation of the meninges, is most often caused by bacteria or viruses. Other pathogens such as fungi or parasites are also causes. Bacterial meningitis is less common than the viral form, can be life-threatening, and is characterized by exudates of polymorphonuclear leukocytes in the CNS. Viral hepatitis is marked mostly by lymphocyte infiltration in the brain and raised numbers of T cells in CSF. A leading cause of meningitis in children is Haemophilus influenzae type b, and a vaccine for it has dramatically reduced its incidence. Meningitis may occur at any age, but it is most common in children, the elderly, and immunocompromised people. 5.4 NEUROCYTOLOGY: CYTOARCHITECTURE The unique cytoarchitecture of the CNS varies regionally. By inspection with the naked eye, the CNS is made up of gray matter and white matter. Unmyelinated neuronal processes and glial cells, tracts of myelinated nerve fibers, and associated glia dominate the white matter, whereas gray matter consists mostly of neuron bodies. The CNS also has a rich vascular supply that includes a profuse network of capillaries, which are more abundant in gray matter. In the spinal cord, gray matter is located internally and is enveloped by an external layer of white matter. In other regions of the CNS, such as cerebrum and cerebellum, an outer cortex of gray matter covers an internal medullary region of white matter. Because of the complexity and intricate nature of nervous tissue, ordinary staining methods have limited value when used alone to examine its cytologic features. 5.5 NEUROCYTOLOGY: STAINING METHODS Special techniques with varied and selective stains, which provide a composite view of nervous tissue, are often used in neurohistology and neuropathology. Basic cationic dyes such as cresyl violet and toluidine blue elucidate cell nuclei and neuronal Nissl substance (rough endoplasmic reticulum and ribosomes). Luxol fast blue and other reagents such as osmium can demonstrate myelin sheaths. Metal impregnation with reduced gold and silver demonstrates the intricate nature of axons and dendrites. Histochemistry and immunocytochemistry permit localization of specific substances and molecules within different types of neurons and glia. Electron microscopy has proved to be quite useful for revealing fine structural details beyond the resolution power of light microscopy. CLINICAL POINT Senile dementia of the Alzheimer type, or Alzheimer disease, which is characterized by progressive memory loss, is increasingly common in developed countries as populations include more elderly persons. Definitive diagnosis is made by microscopic examination of the brain at autopsy, with the histopathologic hallmark being an increased number of neuritic, or senile, plaques, which occur mostly in the cerebral cortex. They consist of tortuous neuritic processes formed mainly by degenerative presynaptic endings, which surround a central amyloid core. Reactive astrocytes and microglia may appear at a plaque periphery. Another important histologic feature is proliferation of intracytoplasmic neurofibrillary tangles. 5.6 STRUCTURE OF A NEURON The neuron is a highly polarized cell that consists of a soma, or cell body, from which cytoplasmic processes arise. The processes, known as nerve fibers, vary greatly in size, some being up to 1.5 m long. Processes conducting impulses toward cell bodies are dendrites, whereas a single process conveying impulses away from cell bodies is an axon. The soma consists of a nucleus and the surrounding cytoplasm, known as the perikaryon. Soma sizes, which depend on cell type and function, vary from 5 to 150 μm. Anterior motor neurons of the spinal cord are among the largest in the CNS, whereas granule cells in the cerebellar cortex are among the smallest. Irregularly shaped masses of basophilic material known as Nissl substance are scattered in the cytoplasm of the body and the dendrites. Neurons are classified into three types on the basis of the number of processes. Multipolar neurons are the most common and characteristic and have one axon and several dendrites. Bipolar neurons have two processes, an axon and a dendrite, and are found in the visual, auditory, and olfactory systems. Pseudounipolar neurons have one short process, which bifurcates into an axon and a dendrite. 5.7 ULTRASTRUCTURE OF A NEURON IN GRAY MATTER IN RELATION TO SURROUNDING STRUCTURES The soma is the trophic center of the neuron and varies greatly in size and shape. Reflecting its role in genetic regulation and transcription, the euchromatic nucleus of a typical neuron has small patches of heterochromatin, peripherally displaced, just under the nuclear envelope. The nucleus is usually spherical to ovoid, and it is large relative to the surrounding perikaryon. A characteristic feature is one or more prominent nucleoli, seen in fortuitous sections; they have a role in synthesizing ribosomal RNA. The surrounding cytoplasm is the site of synthesis of most of the structural and secretory proteins, enzymes, and organelles needed for diverse functions of the cell. Dominating the cytoplasm are multiple flattened cisternae of rough endoplasmic reticulum between which are numerous free ribosomes (the basophilic Nissl substance seen in light micrographs). Their major function is protein synthesis for internal use and export. A highly developed Golgi complex with multiple stacks of flattened sacs and associated vesicles and vacuoles is usually seen near the nucleus. This organelle is responsible for packaging and concentration of secretory products, posttranslational modification of macromolecules, and supply of lysosomes, which accumulate wear-and-tear lipofuscin pigment in these long-lived cells with advancing age. 5.8 ULTRASTRUCTURE OF A SPINAL CORD NEURON SOMA Mitochondria, the source of ATP to meet energy requirements for the metabolically active neuron, are abundant throughout the soma and in peripheral processes of the cell. They are especially numerous in axon terminals close to synapses. Neuronal cytoplasm has a well-developed cytoskeleton consisting of microtubules, actin filaments, and neurofilaments (an intermediate type). These organelles, found throughout the soma and extending into the axon and dendrites, help maintain cell shape and structural stability. Microtubules provide intracellular axoplasmic transport of organelles, most notably of mitochondria and membrane-bound vesicles containing precursors of neurotransmitters. Intracellular neuronal transport is bidirectional: anterograde transport is directed away from the soma and into cytoplasmic processes, whereas retrograde transport transports organelles and other material toward the soma. Microtubules and neurofilaments also play roles in axonal growth and guidance during development and in regeneration after injury. Neurofilaments are best seen by electron microscopy and can be detected by immunocytochemistry, but an affinity for heavy metals is the basis for metal impregnation staining techniques developed more than a century ago. Neuronal cytoplasm also contains lipid droplets, smooth endoplasmic reticulum, lysosomes, peroxisomes, Golgi complexes, ribosomes, and sometimes melanin pigment granules. 5.9 TYPES OF SYNAPSES Synapses are specialized sites for chemical or electrical transmission for communication between neurons or between neurons and other effector cells such as skeletal muscle fibers. Most synapses in humans involve chemical neurotransmitters, which are released from presynaptic terminals of one axon or dendrite to affect receptors on the postsynaptic membrane of the target cell. Various neurotransmitters exist and include amino acids such as glutamate, catecholamines such as epinephrine and norepinephrine, serotonin, neuropeptides, and acetylcholine. In functional terms, two main types of synapses occur: excitatory and inhibitory. In excitatory synapses, neurotransmitter release from the presynaptic neuron depolarizes the postsynaptic membrane; in inhibitory synapses, the postsynaptic membrane is hyperpolarized. Most CNS synapses are between an axon of one neuron and the dendrite of another—axodendritic. Other types include axosomatic and, less commonly, axoaxonic synapses. In some sites, such as hypothalamus and posterior pituitary, large vesicles in presynaptic terminals may contain polypeptide hormones, for example, oxytocin or vasopressin, that are neurosecretory products, not neurotransmitters. 5.10 ULTRASTRUCTURE OF SYNAPSES A typical synapse in the CNS consists of three major components: presynaptic terminal, synaptic cleft, and postsynaptic membrane. The presynaptic terminal aligns closely with the postsynaptic membrane of the target cell. In the area of membrane apposition, presynaptic and postsynaptic membranes are separated by a narrow synaptic cleft 12–30 nm wide. Clusters of large numbers of synaptic vesicles in the presynaptic terminal contain neurotransmitter that is released by exocytosis to mediate synaptic transmission. By electron microscopy, synaptic vesicles are 40–60 nm in diameter and are membrane-bound. Whether they have a clear center or an electron-dense core depends on the chemical nature of the neurotransmitter. Pre- and postsynaptic membrane specializations contain electron-dense material that extends into underlying cytoplasm and is usually thicker in the postsynaptic area. An action potential causes presynaptic vesicles to fuse with the presynaptic membrane and discharge neurotransmitter into the synaptic cleft. Neurotransmitter then diffuses across the cleft to interact with receptor molecules on the postsynaptic membrane, which changes postsynaptic membrane conductance Only gold members can continue reading. Log In or Register to continue Share this: Share on X (Opens in new window) X Share on Facebook (Opens in new window) Facebook Like this:Like Loading… Related Related posts: CARDIOVASCULAR SYSTEM RESPIRATORY SYSTEM SPECIAL SENSES FEMALE REPRODUCTIVE SYSTEM Stay updated, free articles. Join our Telegram channel Join Tags: Netters Essential Histology Jun 18, 2016 | Posted by admin in HISTOLOGY | Comments Off on NERVOUS TISSUE Full access? Get Clinical Tree