CHAPTER 10 Membrane Channels
C hannels are integral membrane proteins with transmembrane pores that allow particular ions or small molecules to cross a lipid bilayer. Some channels are open constitutively, but most open just part time. Each time a channel opens, thousands to millions of ions diffuse down their electrochemical gradient across the membrane. Carriers and pumps are orders of magnitude slower, since they use rate-limiting conformational changes to transport each ion (see Chapters 8 and 9).
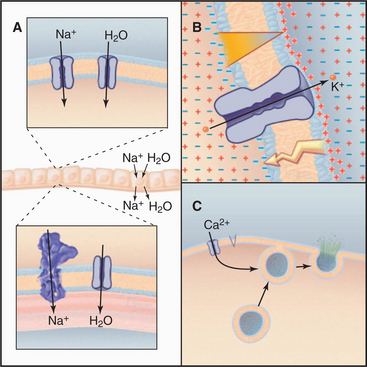
The ability to control diffusion across membranes allows channels to perform three essential functions (Fig. 10-1). First, certain channels cooperate with pumps and carriers to transport water and ions across cell membranes. This is required to regulate cellular volume and for secretion and absorption of fluid, as in salivary glands, kidney, inner ear, and plant guard cells. Second, ion channels regulate the electrical potential across membranes. The sign and magnitude of the membrane potential depend on ion gradients created by pumps and carriers and the relative permeabilities of various channels (Appendix 10-2). Open channels allow unpaired ions to diffuse down concentration gradients across a membrane, separating electrical charges and producing a membrane potential. Coordinated opening and closing of channels change the membrane potential and produce an electrical signal that spreads rapidly over the surface of a cell. Nerve and muscle cells use these action potentials (see Fig. 11-6) for high-speed communication. Third, other channels admit Ca2+ from outside the cell or from the endoplasmic reticulum into the cytoplasm, where it triggers a variety of processes (see Fig. 26-12), including secretion (see Fig. 21-19) and muscle contraction (see Fig. 39-16).
Channels are important in medicine. Ion channels are targets of powerful drugs and toxins, including curare, tetrodotoxin (“voodoo toxin”), paralytic shellfish toxins, cobra toxin, local anesthetics, antiarrhythmic agents, and probably general anesthetics (Table 10-1). Defects in ion channel genes cause many inherited disorders, including some cardiac arrhythmias and kidney stones. In the human autoimmune disorder myasthenia gravis antibodies target ion channels.
Table 10-1 EXAMPLES OF CHANNEL-BLOCKING AGENTS
| Compound (Chemical Class) | Source | Physiological Effect |
|---|---|---|
| Sodium Channel Blockers | ||
| Tetrodotoxin (alkaloid) | Japanese puffer fish | Paralyzes skeletal muscle |
| Saxitoxin (alkaloid) | Dinoflagellates | Paralyzes skeletal muscle |
| μ-Conotoxins (peptide) | Maine snails | Paralyzes skeletal muscle |
| Batrachotoxin (alkaloid) | Arrow poison frogs | Opens Na-channels, paralyzes skeletal muscle |
| Lidocaine | Chemical synthesis | Reduces cardiac and nerve excitability |
| Potassium Channel Blockers | ||
| Quaternary amino alkanes | Chemical synthesis | Blocks K-currents, increases nerve excitability |
| Scorpion toxin | Scorpions | Blocks K-currents, increases nerve excitability |
| Calcium Channel Blockers | ||
| Dihydropyridines | Chemical synthesis | Reduces excitability of L-type channels of striated muscles |
| ω-conotoxin (peptide) | Pacific cone snail | Inhibits nervous system N-type channels; blocks synaptic transmission |
| Nicotinic Acetylcholine Receptor | ||
| α-Bungarotoxin (peptide) | Snake, Bungaris multicinctus | Blocks neuromuscular transmission; paralyzes skeletal muscle |
| α-Cobra toxin | Cobra | Blocks neuromuscular transmission; paralyzes skeletal muscle |
| Curare | Plant, strychnos toxifera | Blocks neuromuscular transmission; paralyzes skeletal muscle |
This chapter covers 12 large families of plasma membrane channels. Other chapters discuss cystic fibrosis transmembrane regulator Cl− channels (see Fig. 11-4), gap junction channels used for communication between adjacent cells (see Fig. 31-6), and intracellular Ca2+ release channels that participate in signal transduction (see Figs. 26-13 and 39-15). Understanding channels requires not only information about their structure and activity but also some knowledge of electrical phenomena. Appendixes 10-1 to 10-3 contain essential material about electrophysiology.
A new channel can be characterized by expressing its cDNA in a test cell and then making electrical recordings of ion currents from the cell or patches of its membrane (Appendix 10-1). If expression of a single-channel protein fails to reproduce the channel activity observed in the cell of origin, auxiliary subunits are probably required. Historically, investigation of channel functions has relied on toxins and drugs that inhibit particular channels more or less specifically (Table 10-1). This approach is often limited by a lack of specificity. Mutations, including those in human disease, provide definitive tests for physiological functions and have yielded some surprising results.
Channel Diversity and Evolution
Humans have about 400 genes that encode channel proteins. The historical channel nomenclature based variously on the ion transported, mode of regulation, physiological role, or drug sensitivity is often ambiguous. Fortunately, knowledge of channel protein structures clarified evolutionary relationships and provided a framework to classify most plasma membrane channels into a few large families (Fig. 10-2).
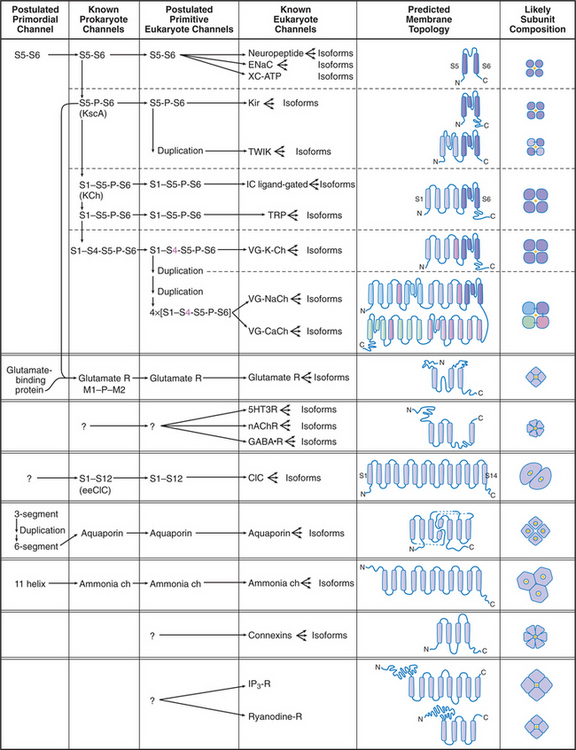
Channels are integral membrane proteins, usually with two or more a-helices crossing the lipid bilayer. Porins are an exception; they are built from transmembrane β-strands (see Fig. 7-8C). Channels generally consist of two to six subunits, but some are single, large polypeptides. The transmembrane pores for conducting ions or other substrates are often located in the middle of a group of subunits or subunit-like domains, but the pores of chloride, water, and ammonia channels are located within single subunits.
Channel Structure
The K+ channel KcsA from the Bacterium Streptomyces lividans serves the model for channels in general and the whole family of S5/S6 channels in particular (Fig. 10-3). Four identical subunits are composed of two transmembrane helices connected by a P loop (for pore)—a short third helix and a crucial strand that makes the selectivity filter. The transmembrane helices are packed close together on the cytoplasmic side of the bilayer but splay apart on the extracellular side to make room for the pore helices and selectivity filter.
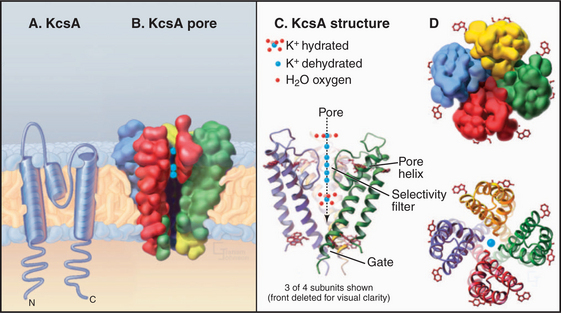
The selectivity filter accommodates two K+ ions, a local concentration exceeding that inside or outside the cell by more than 10-fold, so it actually concentrates K+. However, it does not impede diffusion through the pore, since electrostatic repulsion between these closely spaced ions forces them apart. Outside the filter, the pore is lined with hydrophobic groups, but a cavity in the middle of this passage accommodates a hydrated K+ in an environment with a negative electrostatic potential that is thought to reduce the electrostatic barrier to the ion as it crosses the membrane, as predicted by earlier physiological studies.
Channel Activity
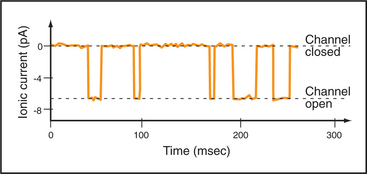
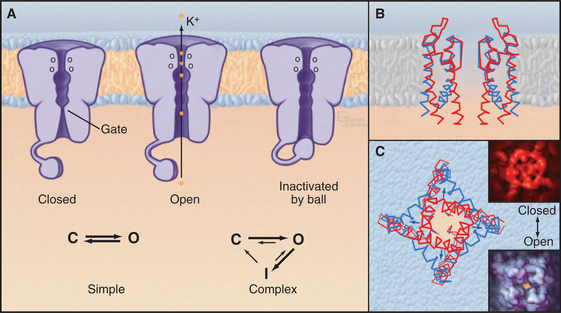
Single-channel electrical recordings show that channel pores are either open or closed (Figs. 10-4 and 10-5). Open channels, also called the active state, pass selected ions across the membrane at rates approaching their diffusion in water. Closed channels have a different conformation that does not pass ions, small solutes, or water. Many channels also have an inactivated state in which part of the channel protein or an impermeant ion blocks the pore of an otherwise open channel, preventing diffusion of ions through the pore. Inactivation makes a channel unresponsive to conditions that favor the active state. Voltage-gated Na+ channels provide a good example; they cycle from closed to open and then inactivate before returning to the closed state.
Selectivity in the Open State
Open channels vary widely in their ability to discriminate among ions. Highly selective channels, such as voltage-gated K+ channels, pass ions without bound water. Less selective channels, such as the nicotinic acetylcholine receptor, are equally permeable to Na+ and K+, which probably pass through as hydrated ions. Gap junction channels pass most molecules smaller than 800 D without discrimination (see Fig. 31-4).
Extensive physiological data and the structure of KcsA suggest that channels achieve their selectivity by virtue of the fact that particular dehydrated ions bind the channel filter as well as their water shell does (Fig. 10-3). Ions that fit poorly in the pore are rejected, as it is energetically unfavorable to shed their hydration shell. The ion flux through an open channel (at a fixed membrane potential) is approximately proportionate to the ion concentration on the side from which the ions migrate. The maximum rate of ion flux—106 to 108 ions/second—is limited by the time required for binding and dissociation at specific sites as an ion traverses the pore. At this high rate, channels discriminate between selected ions that bind and rejected ions that do not during an interaction lasting only 10 to 100 nano-seconds! Ions may move in single file through the pore, driven in part by electrostatic repulsion between the ions.
Transition between the Closed, Open, and Inactivated States
Switching between conducting and nonconducting states is called gating. Gating determines channel activity because channels generally do not open partway or change their ion selectivity. Transitions between closed and open states are so fast that channels are effectively either fully open or fully closed (Fig. 10-4). The steady-state probability of being open (Po) is simply the fraction of the total time that the channel is open. For a given channel, the fraction of time in the open state determines the ion flux. Because channels act independently, the total flux across a membrane depends on the number of channels that are open at a given time.
Comparison of two K+ channel structures shows how a conformational change physically opens and closes a gate (Fig. 10-5). In the closed state (the KcsA structure), the helices at the cytoplasmic end of the pore occlude the lumen. The gate is open in the Ca-gated K+ channel by virtue of a bend in these helices produced by force exerted by a regulatory domain. Energy from Ca2+ binding to the regulatory domain is converted into mechanical work to pull open the gate. Other gating mechanisms are likely to use this principle.
Some channels fluctuate spontaneously between open and closed, but in most cases, local physiological conditions, which are considered in detail in the following sections, control gating from moment to moment. External or internal ligands open some channels. The membrane potential opens and closes other channels without affecting the conductance of the open channels. Mechanical force gates some channels. Cells also use the full range of signaling mechanisms (see Chapters 24 to 26) from phosphorylation to second messengers to guanosine-triphosphate (GTP)–binding proteins to influence the probability that particular channels open or close. By modulating the sensitivity of various channels, cells modify the behavior of their membranes and their responses to external conditions. This modulation makes channels in general, and membrane excitability in particular, highly adaptable. Chapter 11 illustrates how channel modulation regulates the heart rate, changes the efficiency of communication between nerve cells, and adapts cells to some stresses.
In some cases, a process called inactivation stops the flux of ions through active channels. The pore of an inactivated channel remains in the open conformation, so it admits ions, but a part of the channel itself or an ion blocks the pore and prevents ions from crossing the membrane (Fig. 10-5). Flexible cytoplasmic domains inactivate voltage-gated channels (see later discussion) by plugging the open pore. Large organic or inorganic ions, such as polyamines and Mg2+, block other open channels simply by binding within and occluding the pore. The membrane potential influences ion blocking because it drives ions into or out of channels. A blocking ion that binds an open channel and dissociates slowly turns off the channel for a long time. Blockers that dissociate on a millisecond time scale cause the current through the channel to flicker on and off multiple times every time the channel opens. Even faster blocking events cannot be resolved but reduce the rate at which ions move through active channels. Local anesthetics such as lidocaine are pharmacological channel blockers. Binding sites for blocking ions can be found on the outside, the inside, or both sides of the membrane, depending on the channel.
Opening a channel for a few milliseconds can change the membrane potential but not the cytoplasmic ion composition, because only a few ions must cross the membrane to produce a large change in membrane potential (Appendixes 10-2 and 10-3). This conserves energy because ion gradients created by energy-requiring pumps are not dissipated. Longer openings of tens of milliseconds can alter the ion composition of the cell. For example, voltage-gated Ca2+ channels remain open long enough to change the intracellular Ca2+ concentration and trigger cellular events (see Fig. 39-15 B). In this way, they convert an electrical signal to a chemical signal.
Channels with One Transmembrane Segment
The simplest known channel is found in the membrane envelope of influenza virus. This M2 channel consists of four small subunits, each with but one transmembrane helix. After an infected cell takes the virus into an endosome (see Chapter 22), the acidic environment opens the channel, allowing protons to enter the virus and to begin to disassemble the protein shell that surrounds the genome. The antiviral drug amantadine blocks these channels. Bacteria secrete peptides (gramicidin, alamethicin, and colicins) that are designed to kill other species by forming highly selective and conductive channels. Only 13 amino acids are required for gramicidin A to form β-helical homodimers that function as K+-selective channels.
Channels with Two Transmembrane Segments
Mechanosensitive Channels
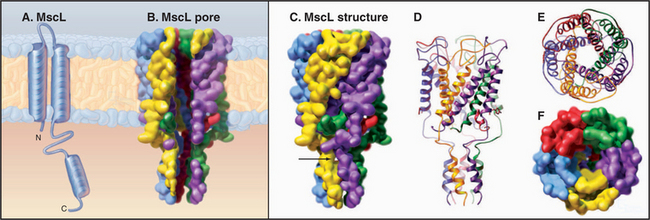
MscL from Mycobacterium tuberculosis (Fig. 10-6) is a simple channel of five subunits. One of the two transmembrane helices forms the wall of the pore. A third C-terminal helix extends the pore 4 nm into the cytoplasm. The central pore is lined with polar residues except for a gate at its narrowest constriction, where an isoleucine and a value reduce the diameter to about 0.2 nm. Tension in the plane of the membrane created by osmotic stress is believed to rearrange these helices and open the channel. Cations pass through the open channels indiscriminately at high rates, since they lack a selectivity filter like that of KcsA. This response avoids osmotic lysis of the cell. Such channels are widespread in prokaryotes and are also found in eukaryotes.
Inward Rectifier Potassium Channels
The Kir family of channels has the same evolutionary origin as KcsA (S5-P-S6 [Fig. 10-2]). Like KcsA (Fig. 10-3), Kir channels consist of two transmembrane helices with a P loop in between. The P loop and helix S6 line the K+-selective pore. In spite of these common features, these channels vary in many respects.
Several channels in this family (Kir2.1, Kir2.3, Kir3 family, and Kir4.1) are inward rectifiers. A rectifier is an electronic component that passes current preferentially in one direction. Inward rectifier K+ channels pass K+ into the cell when the membrane potential is below EK (Appendix 10-2), a membrane potential that is not achieved physiologically. Above the resting potential, these K+ channels pass only a small K+ current out of the cell when they open. The reason is that impermeant cytoplasmic cations, Mg2+, and polyamines (ornithine metabolites having net positive charges of 2+ to 4+) bind to negatively charged residues on the cytoplasmic end of the S6 segment of open channels and block the passage of K+. Despite their low permeability, these channels help to maintain the resting membrane potential in many cells and to repolarize excitable cells during an action potential.
Epithelial Sodium Channels
Epithelial Na+ channels accelerate the rate-limiting step in Na+ transport, an essential process that moves salt and water across epithelia in a number of organs (Fig. 10-1A). Typically, epithelial Na+ channels in the apical plasma membrane provide pores for Na+ to diffuse down its concentration gradient into the cytoplasm, and Na+/K+-ATPases in the basolateral plasma membrane pump Na+ out of the cell into the underlying extracellular space. Water follows Na+ through water channels. Renal collecting tubules use this strategy to resorb salt and water. Lung epithelial cells do the same to clear fluid from air spaces. Mice with knockout mutations in the lung epithelial Na+ channel gene die at birth with fluid in their lungs.
Epithelial Na+ channels consist of multiple α-, β-, and γ-subunits, but their stoichiometry is not known. All have two hydrophobic segments that are thought to be transmembrane helices but no verified P loops. The second putative helix probably lines a pore that is 10 times more permeable to Na+ than to K+. The channel opens and closes randomly for relatively long periods, between 0.5 and 5 seconds, unaffected by the membrane potential or any known natural ligand. The drug amiloride blocks epithelial Na+ channels, so they are called amiloride-sensitive Na+ channels to distinguish them from voltage-gated Na+ channels. The steroid hormone aldosterone, produced in response to salt loss, increases the plasma membrane content of Na+ channels and the rate of Na+ resorption by the kidney.
Channels with Four Transmembrane Helices
K+ channels with four transmembrane segments and two P loops (Fig. 10-2, TWIK) are abundant in animal genomes, with 40 to 50 genes in C. elegans. Two of these subunits form a channel with four domains similar to KcsA. They help to establish the resting potential of the plasma membrane by allowing K+ to leak out of the cell, independent of the membrane potential. These leak channels are activated by volatile anesthetics, leading to hyperpolarization of the membrane and reduced excitability.
Voltage-Gated Cation Channels
Voltage-gated channels have two main functions. First, voltage-gated K+ and Na+ channels produce action potentials in excitable cells (see Fig. 11-6). Depolarization of the membrane opens these channels transiently, driving the membrane potential first toward the Na+ equilibrium potential (Appendix 10-2) and then back toward the K+ equilibrium potential. Second, voltage-gated Ca2+ channels convert electrical signals into chemical signals when they admit Ca2+ to the cytoplasm, where it acts as a second messenger (see Figs. 11-8, 11-9, and 26-12) to stimulate secretion, activate protein kinases, trigger muscle contraction, or influence gene expression.
Voltage-gated channels share a common domain organization (Fig. 10-2). Crystal structures of voltage-gated K+ channels from a thermophilic Archaea and rat brain (Fig. 10-7
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


