CHAPTER 30 Cellular Adhesion
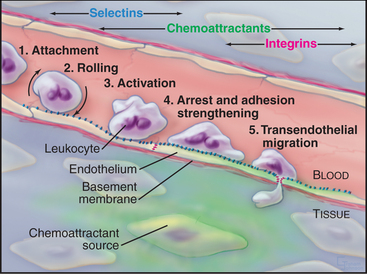
All cells interact with molecules in their environment, in many cases relying on cell surface adhesion proteins to bind these molecules. Multicellular organisms are particularly dependent on adhesion of cells to each other and the extracellular matrix (ECM). During development, carefully regulated genetic programs specify cell-cell and cell-matrix interactions that determine the architecture of each tissue and organ. Some adhesive interactions are stable. Muscle cells must adhere firmly to each other and to the connective tissue of tendons to transmit force to the skeleton (see Chapter 39). Skin cells must also bind tightly to each other and the underlying connective tissue to resist abrasion (see Fig. 35-6). On the other hand, many cellular interactions are transient and delicate. At sites of inflammation, leukocytes bind transiently to endothelial cells lining small blood vessels and then use transient interactions with the ECM to migrate through connective tissue (look ahead to Figs. 30-13 and 30-14).
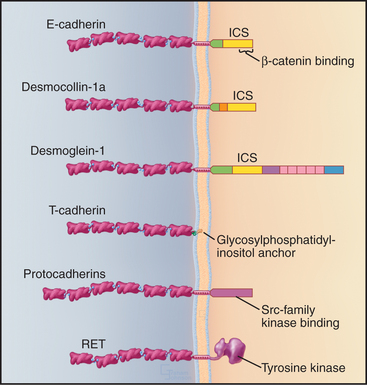
Cells use a relatively small repertoire of adhesion mechanisms to interact with matrix molecules and each other. This conceptual breakthrough came when comparisons of amino acid sequences showed that most adhesion proteins fall into five large families (Fig. 30-1). Within each of these distinctive families, ancestral genes duplicated and diverged during evolution, giving rise to adhesion proteins with the many different specificities that are required for embryonic development, maintenance of organ structure, and migrations of cells of our defense systems. Common properties within each family allowed the appreciation of general mechanisms to emerge from characterizing a few examples. Several important adhesion proteins fall outside the five major families, and additional families may emerge from continued research.
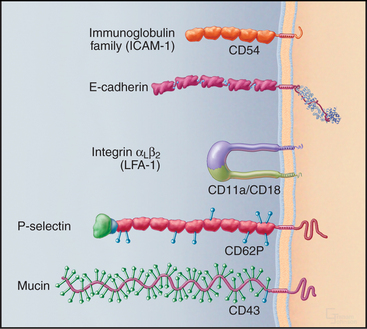
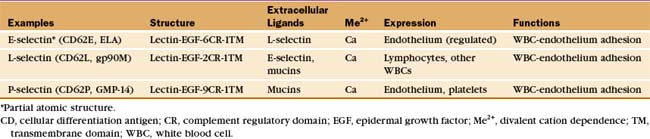
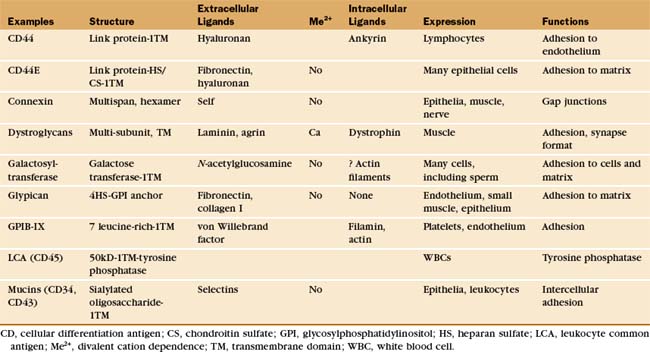
Many adhesion proteins were named before they were classified into families. Tables 30-1 through 30-5 are designed to help the reader with the nomenclature. Many adhesion proteins are named “CD” followed by a number. This stands for “clusters of differentiation,” a term that is used to classify cell surface antigens recognized by monoclonal antibodies, independent of any knowledge about the structure or function of the antigen. Hence, members of the four major families of adhesion proteins have CD numbers.
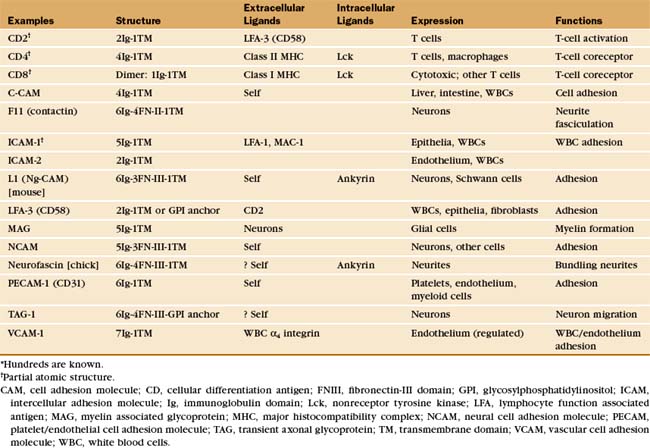
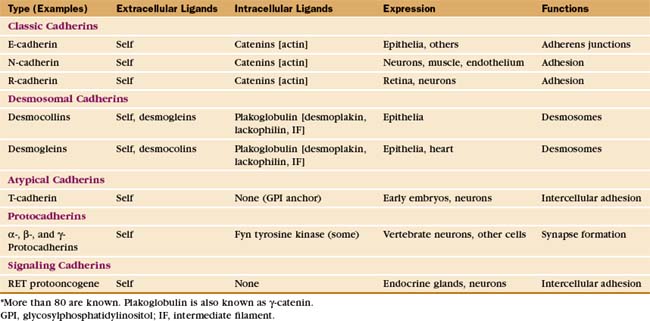
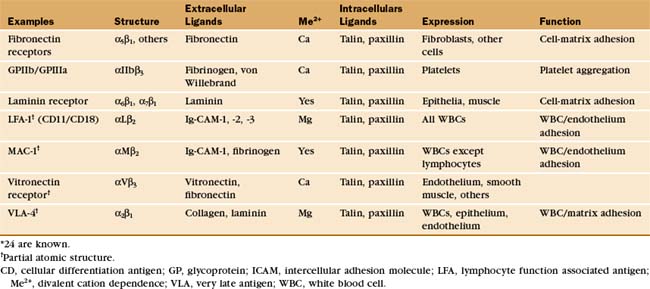
This chapter first highlights some general features of adhesion proteins and then introduces four major families: immunoglobulin–cell adhesion molecules (Ig-CAMs), cadherins, integrins, and selectins. While learning about each family, the reader should not lose track of an important point: These receptors rarely act alone. Rather, they usually function as parts of multicomponent systems. Two examples at the end of the chapter illustrate the cooperation that is required for leukocytes to respond to inflammation and for platelets to repair damage to blood vessels. Chapter 31 on intercellular junctions, Chapter 32 on specialized connective tissues, and Chapter 38 on cellular motility provide more examples of cellular adhesion.
General Principles of Cellular Adhesion
Second Principle of Adhesion
Many adhesion proteins bind one main ligand, and many ligands bind a single type of receptor (refer to Tables 30-1 through 30-5). If this one-to-one pairing were the rule, adhesion would be simple indeed. However, many exceptions exist, particularly in the integrin family of receptors (Table 30-3). These receptors generally bind more than one ligand, and some ligands, such as fibronectin, bind more than one integrin. One can generalize about the ligands for the several families of cell adhesion molecules:
Third Principle of Adhesion
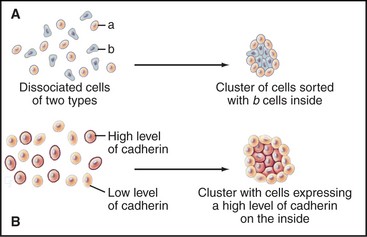
Cells modulate adhesion by controlling the surface density, state of aggregation, and state of activation of their adhesion receptors. Surface density reflects not only the level of synthesis but also the partitioning of adhesion molecules between the plasma membrane and intracellular storage compartments. For example, endothelial cells express P-selectin constitutively but store it internally in membranes of cytoplasmic vesicles. When inflammatory cytokines activate endothelial cells, these vesicles fuse with the plasma membrane, exposing P-selectin on the cell surface, where it binds white blood cells (Fig. 30-13). The importance of surface density is illustrated by an experiment in which cells that express different levels of the same cadherin are mixed together. Over time, they sort out from each other, the more adherent cells forming a cluster surrounded by the less adherent cells (Fig. 30-2). Such differential expression of cadherin determines the position of the oocyte in Drosophila egg follicles. Intracellular signals control the extracellular binding activity of integrins and cadherins. A variety of extracellular stimuli activate intracellular signaling pathways in lymphocytes, platelets, and other cells, which enhance or inhibit the ligand-binding activity of integrins already located on the cell surface. Integrin activation also regulates cellular interactions during development.
Fourth Principle of Adhesion
The rates of ligand binding and dissociation are important determinants of cellular adhesion. Many cell surface adhesion proteins (including members of the Ig-CAM, cadherin, integrin, and selectin families) bind their ligands weakly in comparison with other specific macromolecular interactions, such as the interaction of antigens and antibodies, hormones and receptors, or transcription factors and DNA. The measured dissociation equilibrium constants for these adhesion receptors are in the range of 1 to 100 mM, reflecting high rate constants (>1 s−1) for dissociation of ligand. In some cases, this makes good biological sense. Rapidly reversible interactions allow white blood cells to roll along the endothelium of blood vessels (Fig. 30-13). Transient adhesion also allows fibroblasts to migrate through connective tissue. On the other hand, the interactions of cells in epithelia and muscle appear to be more stable, perhaps owing to multiple weak interactions between clustered adhesion proteins cooperating to stabilize adherens junctions and desmosomes (Fig. 31-7). The combined strength of these bonds is said to increase the “avidity” of the interaction.
Identification and Characterization of Adhesion Receptors
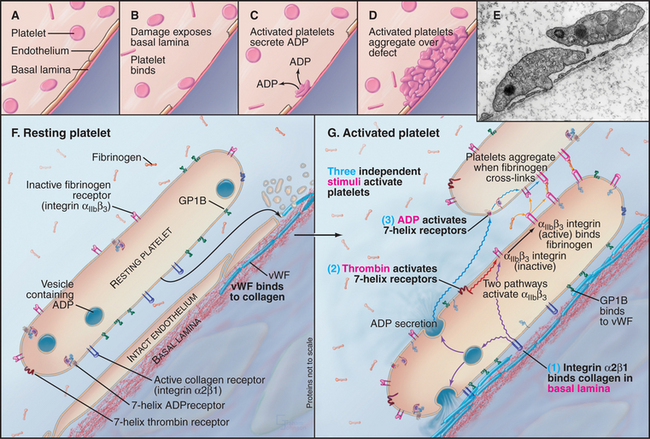
Insights about the functions of adhesion receptors have usually come in several steps. Localization of a protein on specific cells frequently provides the first clues. Typically, the expression of each protein is restricted to a subset of cells or to a specific time during embryonic development or both. Next, investigators use specific antibodies to test for the participation of the adhesion protein in cellular interactions in vitro or in tissues. Blistering skin diseases called pemphigus illustrate the serious consequences when pathological autoantibodies disrupt adhesion between skin cells expressing the antigen (see the sections “Desmosomes” and “Adhesion to the Extracellular Matrix” in Chapter 31). Both human genetic diseases and experimental genetic knockouts in mice and other organisms produce defects caused by the absence of adhesion proteins. In leukocyte adhesion deficiency, white blood cells lack the b2 integrin that is required to bind the endothelial cells that line blood vessels. These defective white blood cells fail to bind to blood vessel walls or to migrate into connective tissue at sites of infection. Similarly, patients with a bleeding disorder called Bernard-Soulier syndrome lack one of the adhesion receptors for von Willebrand factor, a protein that promotes platelet aggregation. Loss of cadherins contributes to the spread of some cancer cells.
Immunoglobulin Family of Cell Adhesion Molecules
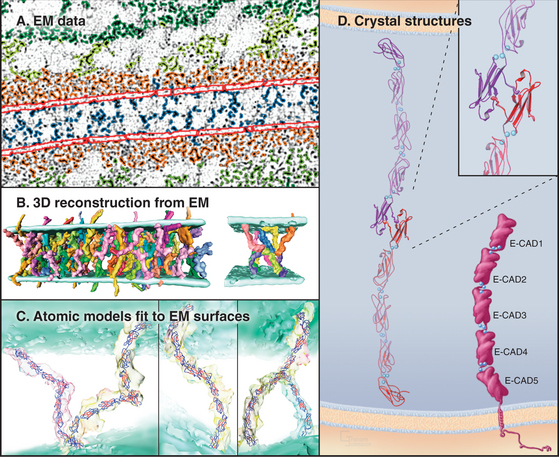
The Ig-CAM family contains hundreds of adhesion proteins, each with one to seven extracellular domains, similar to immunoglobulin domains, anchored to the plasma membrane by a single transmembrane helix (Fig. 30-3 and Table 30-1). Crystal structures established the antibody-like fold of the extracellular domains of several Ig-CAMS. These compact Ig domains consist of 90 to 115 residues folded into seven to nine β-strands in two sheets, usually stabilized by an intramolecular disulfide bond. The N- and C-termini are at opposite ends of these domains, allowing the formation of linear arrays of immunoglobulin domains.
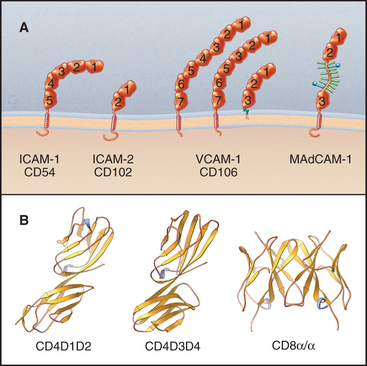
Some Ig-CAMs consist of a single polypeptide, but others are multimeric, with two (CD8) or four (see Fig. 27-8 for the T-cell receptor) subunits. Some nervous system Ig-CAMs have three or four fibronectin III (FN-III) domains between the immunoglobulin domains and the membrane anchor. The C-terminal cytoplasmic tails of these receptors vary in sequence and binding sites. The cytoplasmic domains of the lymphocyte accessory receptors CD4 and CD8 bind protein tyrosine kinases required for cellular activation (see Fig. 27-8). The cytoplasmic domains of neuronal Ig-CAMs bind PDZ domain proteins or membrane skeleton (see Fig. 7-10).
Differentiated metazoan cells express Ig-CAMs selectively, especially during embryonic development, when they may contribute to the specificity of cellular interactions required to form the organs. Neurons and glial cells express specific Ig-CAMs that guide the growth of neurites, mediate synapse formation and promote the formation of myelin sheaths. In adults, interaction of endothelial cell ICAM-1 with a white blood cell integrin is essential for adhesion and movement of the leukocytes into the connective tissue at sites of inflammation (Fig. 30-13).
Like other cell adhesion proteins, Ig-CAMs participate in signaling processes. Best understood are interactions of lymphocytes with antigen-presenting cells during immune responses. Ig-CAMs reinforce the interaction of antigen-specific T-cell receptors with major histocompatibility complex molecules carrying appropriate antigens on other cells (see Fig. 27-8). Although individual interactions are weak, the combination of specific (T-cell receptor) and nonspecific (CD2 and CD4) interactions with the target cell is sufficient to initiate signaling.
Cadherin Family of Adhesion Receptors
The complex architecture of organs in vertebrates depends on Ca2+-dependent associations between the cells mediated by more than 80 cadherins (Table 30-2). Their name derives from “calcium-dependent adhesion” protein. Genes for cadherin domains appeared in unicellular precursors of sponges, an early step toward the evolution of metazoan organisms.
Cadherins generally interact with like cadherins on the surfaces of other cells in a calcium-dependent fashion, but research is uncovering a growing list of examples of heterophilic interactions. Homophilic interactions of cadherins link epithelial and muscle cells to their neighbors, especially at specialized adhesive junctions called adherens junctions and desmosomes (Fig. 30-4; also see Fig. 31-7). The cytoplasmic domains of cadherins interact with actin filaments or intermediate filaments to reinforce these junctions and maintain the physical integrity of tissues. Contacts mediated by cadherins also influence cellular growth and migration, including suppression of growth and invasion of tu-mors, as well as formation of synapses in the nervous system.
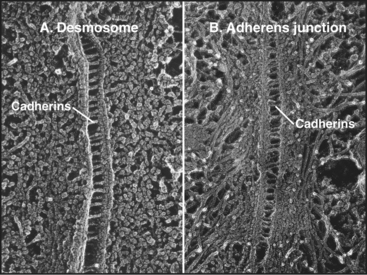
The structural hallmark of the cadherin family is the CAD domain (Figs. 30-5 and 30-6). CAD domains consist of about 110 residues folded into a sandwich of seven β-strands. This fold is similar to immunoglobulin and FN-III domains, but the limited sequence homology suggests independent origins and convergent evolution. N- and C-termini are on opposite ends of CAD domains. Ca2+ bound to three sites between adjacent CAD domains links them together into rigid rods. Without Ca2+, the domains rotate freely around their linker peptides.