CHAPTER 45 Meiosis*
Meiosis (from the Greek, meaning “reduction”) is a specialized program of two coupled cell divisions used by eukaryotes to maintain the proper chromosome number for the species during sexual reproduction. The number of chromosomes is halved in meiosis; therefore, the subsequent fusion of male and female gametes restores the proper chromosome number for the species. The reduction in chromosome number is achieved by randomly separating homologous chromosomes, each pair of which is composed of one chromosome donated by the mother and one donated by the father. This pairing and subsequent separation of homologous chromosomes are typically made possible by genetic recombination, which occurs during the lengthy and complex prophase of the first meiotic division. The random segregation of homologous chromosomes and the genetic recombination that make this possible form the physical basis of the laws of classical genetics, first proposed by Gregor Mendel in 1866.
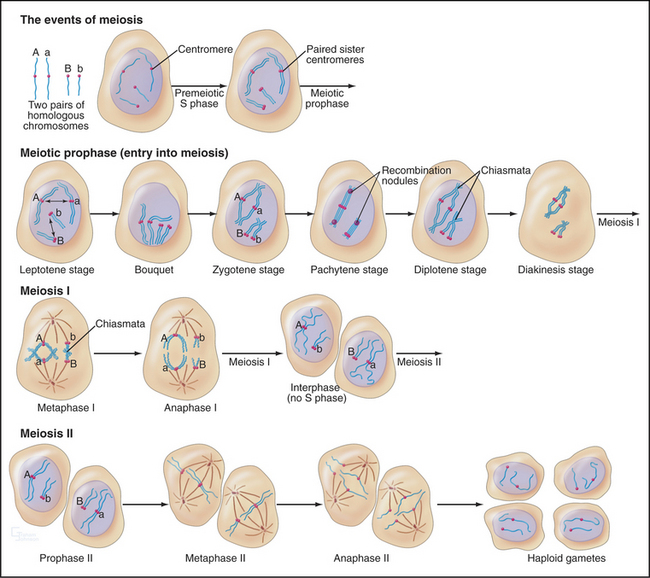
The unique events of meiosis occur in the first division, termed meiosis I (Figs. 45-1 and 45-2). Because the daughter cells have half the number of chromosomes, meiosis I is also known as the reductional division. The second division, meiosis II, is similar in most respects to mitosis: Sister chromatids segregate from each other, and the number of chromosomes remains the same (Box 45-1; see also Chapter 44). Meiosis II is called the equational division. Meiosis is an ancient process that occurs in virtually all higher eukaryotes, including the animal, fungal, and plant kingdoms.
BOX 45-1 Important Differences between Meiosis and Mitosis
The products of meiosis are haploid. The products of mitosis are diploid.
Meiosis: An Essential Process for Sexual Reproduction
Without meiosis, there would be no sex because every fusion of gametes would increase the number of chromosomes in the progeny. Sexual reproduction is an important survival strategy that offers organisms a mechanism for altering the genetic makeup of offspring. This strategy has been conserved throughout higher eukaryotes and is inextricably linked with the mechanism of meiosis.
Meiosis I also produces novel versions of chromosomes by exchange of DNA segments between homologs. This occurs because each chromosome must typically undergo at least one genetic recombination (crossover) event to segregate properly at anaphase of meiosis I. If the chromosomes of all the individuals of a species were identical, meiosis and sexual reproduction would only provide different combinations of the same chromosomes. However, human chromosomes vary between individuals, averaging about one difference (polymorphism) per 1000 base pairs, and it is estimated that, overall, at least 106 sites across the genome have variant versions. Recombination involves exchange of chromosomal segments, producing new chromosomes that are a patchwork of segments from the maternal and paternal homologs. The combined effects of recombination and random assortment of homologs in meiosis I yields a vast number of different gametes and provides an important source of genetic diversity that permits eukaryotic populations to adapt to changing environmental conditions.
The Language of Meiosis
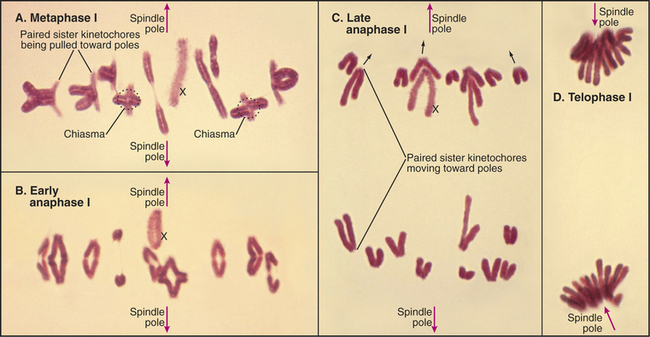
Meiosis is all about the segregation of the paired homologous chromosomes. This process shows some key differences from mitosis (Box 45-1). When the homologs are balanced at the metaphase plate of the meiosis I spindle, it is the chiasmata that hold them together and counteract the pulling force of the spindle on the kinetochores (Fig. 45-2). Cohesion between the chromatid arms holds chiasmata in place until it is released at anaphase of meiosis I. Centromeres of the sister chromatids remain associated with one another throughout meiosis I until anaphase of meiosis II. This means that at anaphase, when the chiasmata are released, each pair of sister chromatids migrates to the same spindle pole. As a result, the progeny of meiosis I have the haploid number of chromosomes each paired with a sister chromatid. Box 45-2 reviews some genetics terms that are helpful in understanding meiosis.
BOX 45-2 Brief Overview of Genetic Terminology
A comprehensive introduction to the field of genetics is beyond the scope of this text. However, here are a number of terms used by geneticists that will assist in the understanding of the discussion of genetic recombination and its role in meiosis (also see Box 6-2).
Two types of recombination events occur during meiosis (Fig. 45-3). The first of these—noncrossover events (frequently referred to as gene conversion)—may involve the loss of one or more genetic markers. Noncrossover events are the most common outcome of the programmed double-strand DNA breaks that occur during leptotene. They are thought to involve the invasion of a double helix by a region of single-stranded DNA with complementary sequence but then ejection of this sequence before assembly of a Holliday junction and completion of recom-bination.
Recombination
Because recombination is the key to the behavior of chromosomes in meiosis I, this process is discussed in detail herein to provide a mechanistic underpinning for understanding later events. Meiotic recombination is very similar to the process of homologous recombinational repair of double-strand DNA breaks in somatic cells (review Box 43-1 and Fig. 43-15 as a prelude to studying meiotic recombination).
Spo11 generates programmed double-strand DNA breaks very early during meiotic prophase (Fig. 45-3). Spo11 is a type II DNA topoisomerase (see Chapter 13, under the section titled “Proteins of the Mitotic Chromosome and Chromosome Scaffold”) that cleaves both DNA strands in a reaction that produces a covalent linkage between a tyrosine on the enzyme and the cleaved phosphodiester backbone. Where it has been measured, Spo11 creates about threefold to fivefold more DNA breaks than ultimately complete the recombination pathway to produce reciprocal exchanges of DNA between homologous chromosomes, or crossovers. An alternative pathway is thought to process the excess breaks, producing noncrossover events (Box 45-2 and Fig. 45-3I-J). Each pair of homologous chromosomes thus undergoes many noncrossover events and a very few crossover events (often only one) during meiosis I prophase.
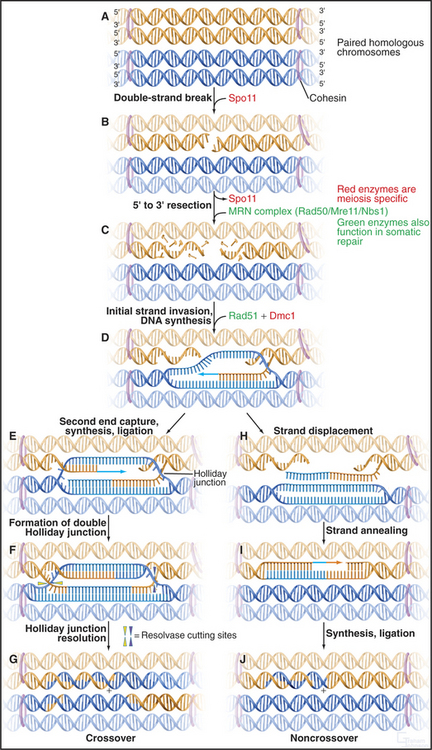
Figure 45-3 the events of recombination. Recombination occurs between homologs rather than sisters. A, Paired homologous chromosomes. Sister chromatids are held tightly together by cohesin, shown here schematically as hoops. B, Spo11 makes a double-strand break. C, Resection of the break. D, First strand invasion. At this point, the pathway splits in two, one outcome leading to a crossover and the other to a noncrossover. Crossover pathway: E, The second resected strand invades its homologous partner. New DNA synthesis fills the gaps. F, The resulting molecule contains a double Holliday junction (see Fig. 43-15B). If the resolvase (nuclease) cuts the double Holliday junction asymmetrically as shown (i.e., one vertical and one horizontal cut), the result is a crossover (G). If the cuts are symmetrical, a noncrossover molecule is produced. Noncrossover pathway: H, In most cases, the invading DNA strand is ejected prior to stabilization and formation of a double Holliday junction. I, DNA gap-filling and ligation yield a noncrossover chromosome (J).
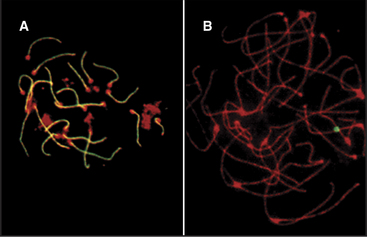
In both mice and yeast, double-strand breaks generated by Spo11 are required for normal segregation of homologous chromosomes. In Spo11-null mice, recombination is not initiated, and synapsis, if it occurs at all, is aberrant, often involving nonhomologous chromosomes (Fig. 45-4). In these mutant mice, spermatocytes die by apoptosis early in meiotic prophase, and oocytes die somewhat later. In contrast, the nematode Caenorhabditis elegans and the fruit fly Drosophila melanogaster do not require Spo11-induced double-strand breaks for synapsis of homologous chromosomes.
Once the DNA double-strand breaks have been produced, they are processed by the 5′ → 3′ exonuclease MRN (Mre11/Rad50/Nbs1), which chews back one strand of the double helix (a process called resection), leaving single-stranded tails at the 3′ end of the DNA molecules (Fig. 45-3C; see also Fig. 43-15). The same exonuclease functions in somatic DNA repair and in meiotic recombination.
Next, the single-stranded tails “invade” the other chromosomes, looking for complementary DNA se-quences. This process is driven by Rad51 and Dmc1, two proteins that are related to the E. coli RecA protein, which is essential for DNA recombination in bacteria. These proteins polymerize into nucleoprotein filaments on DNA and use ATP hydrolysis to catalyze homologous pairing and strand exchange reactions. The process inserts a single-stranded region of DNA into a double helix, displacing one of the two paired strands. Dmc1 functions only in meiosis, but Rad51 has other essential functions as well. Dmc1 may promote the search for homologous chromosomes, rather than sister chromatids as occurs in somatic DNA repair. Mutants that lack Dmc1 are defective in homologous chromosome pairing. Rad51p and Dmc1p are found in structures called early recombination nodules that are distributed along the chromosome axes early in meiosis (Fig. 45-9).
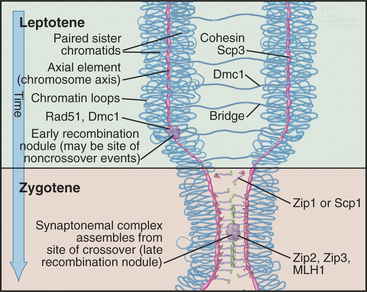
It is now believed that if only one single strand successfully invades the homologous chromosome, the outcome is a noncrossover event, whereas invasion of both single-strand tails leads to crossovers. The double invasion produces branched intermediates known as double Holliday junctions (Fig. 45-3F-G; see also Fig. 43-15B). These are then cleaved by as yet unknown nucleases and converted to mature crossover recombination products.
Tracking the Homologous Chromosomes through the Stages of Meiotic Prophase I
Pairing and recombination of homologous chromosomes take place during prophase of meiosis I. In the discussion of these processes, it is necessary to refer to the five stages of meiotic prophase: leptotene, zygotene, pachytene, diplotene, and diakinesis (Fig. 45-1). As the understanding of meiotic prophase advances, the significance of these stages is being reassessed. In particular, these morphologic stages do not correspond directly to the steps of meiotic recombination, as was assumed previously.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


