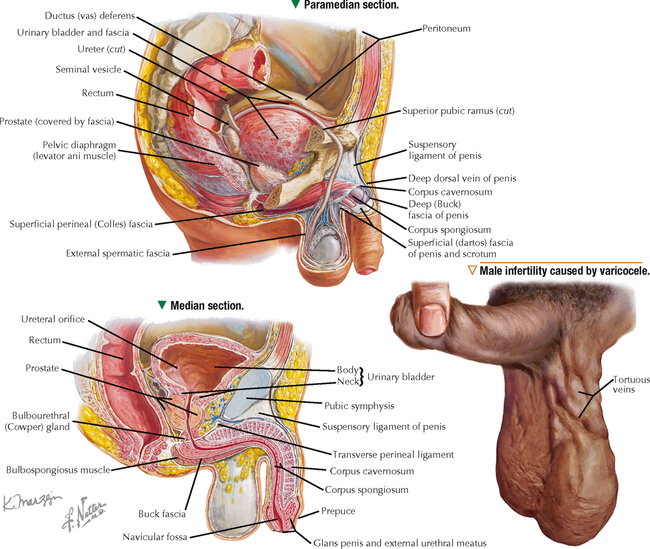
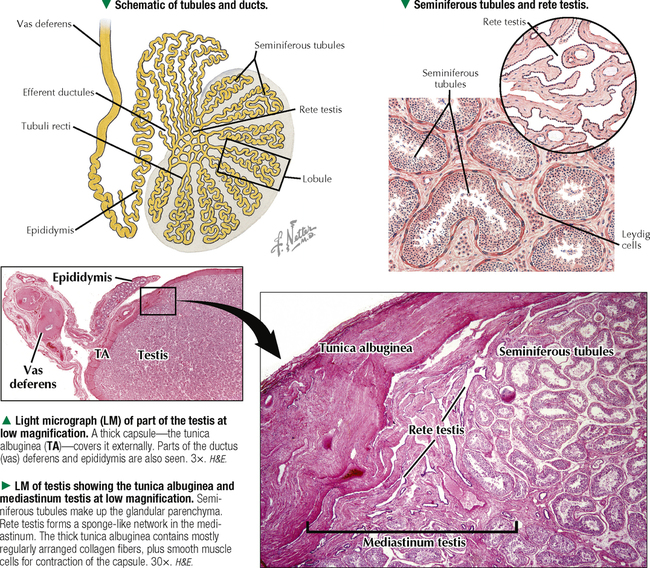
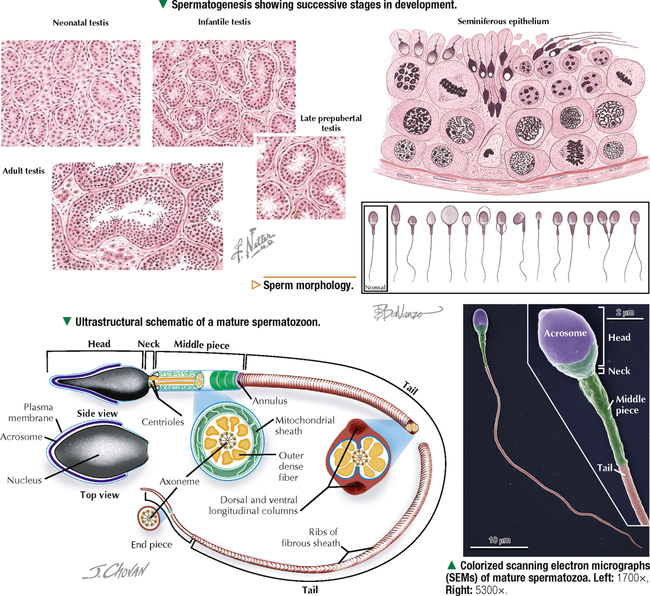
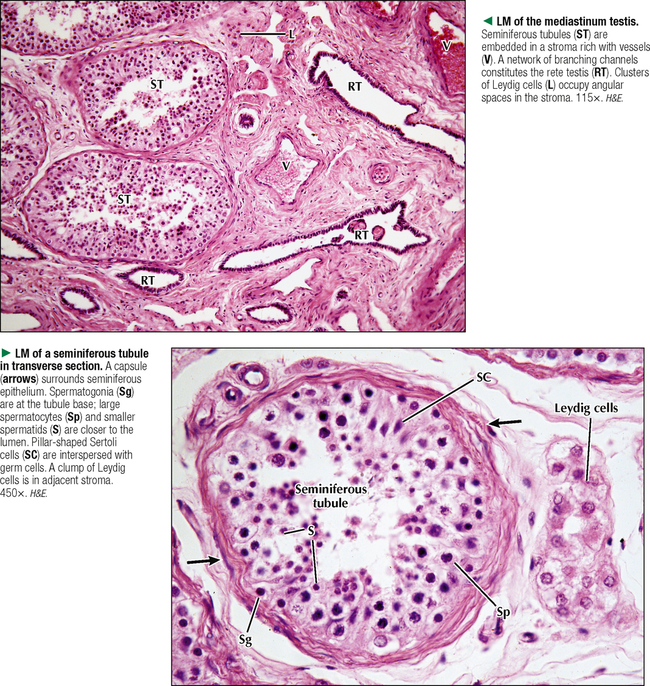
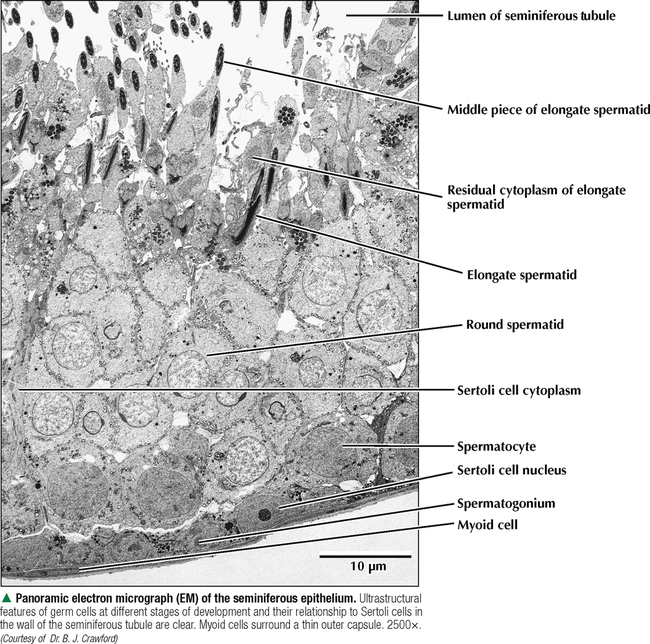
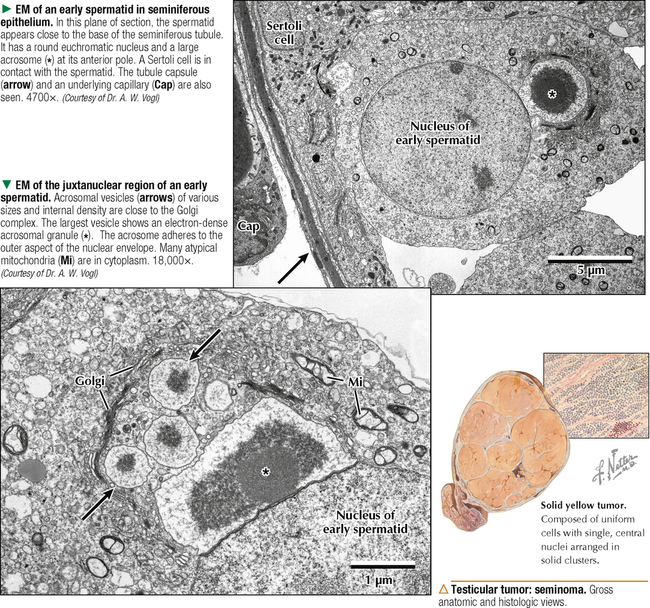
17 MALE REPRODUCTIVE SYSTEM 17.1. Overview 17.2. Anatomy and Histology of Testes 17.3. Testicular Development and Spermatogenesis 17.4. Histology of Seminiferous Tubules 17.5. Ultrastructure of Seminiferous Tubules and Spermatogenesis 17.6. Ultrastructure of Germ Cells and Early Spermiogenesis 17.7. Ultrastructure of Germ Cells and Later Spermiogenesis 17.8. Ultrastructure and Function of Sertoli Cells 17.9. Histology and Ultrastructure of Leydig Cells 17.10. Ultrastructure and Function of Leydig Cells 17.11. Anatomy and Histology of the Epididymis 17.12. Histology and Function of the Epididymis 17.13. Histology of the Ductus (Vas) Deferens 17.14. Ultrastructure and Function of the Ductus (Vas) Deferens 17.15. Anatomy and Histology of the Prostate and Seminal Vesicles 17.16. Histology and Function of the Prostate 17.17. Ultrastructure of the Prostate 17.18. Histology of Seminal Vesicles 17.19. Anatomy and Histology of the Urethra and Penis 17.20. Histology of the Penis 17.21. Histology and Function of the Penis 17.1 OVERVIEW The male reproductive system includes the paired primary sex organs, the testes, which have both exocrine and endocrine functions, and several secondary sex organs consisting of excretory ducts and accessory glands. The scrotum and penis, an erectile organ through which the distal urethra passes, are external genitalia. The testes reside outside the body cavity in the scrotum, where they are suspended and maintained in position. Testes and associated spermatic cords are invested by distinct layers of tissue acquired during descent of male gonads from their original retroperitoneal position in the abdominal cavity to the scrotum. Testes contain small convoluted seminiferous tubules, whose germinal epithelium produces male germ cells known as spermatozoa, and interstitial connective tissue. Leydig (interstitial) cells produce testosterone, the hormone responsible for male secondary sex characteristics. After spermatozoa are produced in the testes, they travel a long, tortuous route: from seminiferous tubules to the paired rete testis, efferent ductules (ductuli efferenti), epididymis, ductus (vas) deferens, and ejaculatory ducts, and to the single urethra and penis. The accessory glands include two seminal vesicles, single prostate gland, and paired bulbourethral glands that secrete into ejaculatory ducts and urethra. Three key functions of this system are production of spermatozoa, delivery of these cells via semen into the female reproductive tract, and production of testosterone. CLINICAL POINT The most common causes of male infertility—failure to initiate a pregnancy in a fertile female partner—are a low sperm count, production of immobile or misshapen spermatozoa, and blockages along the male reproductive tract that prevent normal delivery of semen. Other causes include testicular damage caused by cryptorchism (undescended testes), orchitis (inflammation of testes), and varicocele (scrotal varicose veins). Radiation, excessive tobacco use or alcohol consumption, and hormonal disturbances may also contribute to it. 17.2 ANATOMY AND HISTOLOGY OF TESTES The testis, an ovoid gland weighing about 15 g, is encased in a thick capsule of dense fibroelastic connective tissue known as the tunica albuginea, because it appears white in life. An outer visceral layer of the tunica vaginalis invests the capsule externally. Along the posterior border of the testis, the capsule projects inward as a thickened ridge, known as the mediastinum testis. This ridge corresponds to the hilum of other glands—the site where ducts, blood vessels, lymphatics, and nerves connect to the gland interior. Thin fibrous partitions, or septa, radiate from the mediastinum and form wedge-shaped lobules, about 250 in the human. The lobules contain seminiferous tubules, which are sectioned in different planes because they have a convoluted course. Each testis has 600–1200 seminiferous tubules, with a total length of 280–400 m. In the mediastinum, seminiferous tubules empty into tubuli recti and rete testis, which coalesce to form six to eight efferent ductules. These ducts drain testicular fluid and spermatozoa to the proximal part of the epididymis. The rete testis is a labyrinthine network of collecting chambers of simple cuboidal epithelium. Interstitial connective tissue constitutes 20%-30% of the substance of the gland and consists of vascularized connective tissue with clusters of hormone-producing Leydig cells. CLINICAL POINT Orchitis—inflammation of one or both testes—is usually caused by a viral, bacterial, or fungal infection. Viral orchitis, most commonly caused by the mumps virus, typically affects boys after puberty. Most cases of bacterial orchitis occur in young men via sexually transmitted pathogens (e.g., gonorrhea, trachomatis, Chlamydia) or in older men with benign prostatic hypertrophy. Orchitis may also accompany infections spread from other regions (e.g., epididymitis, prostatitis, cystitis). The diagnosis is via urinalysis, urine culture, complete blood count (CBC), and testicular ultrasonography. 17.3 TESTICULAR DEVELOPMENT AND SPERMATOGENESIS Testes derive from embryonic intermediate mesoderm that initially gives rise to primary epithelial sex cords, the precursors of seminiferous tubules. In the 4-week embryo, primordial germ cells migrate from yolk sac endoderm to the cords. Newborn testis consists of solid cords of germ cells arranged in layers and closely associated with epithelial cells that will be supporting Sertoli cells. The cords remain solid until puberty, when they lengthen, increase in diameter, and acquire a lumen. Leydig cells develop from mesenchyme between the seminiferous tubules. At puberty, primitive germ cells—spermatogonia—enlarge and become mitotically active. These cells undergo the process of spermatogenesis, in which diploid spermatogonia in seminiferous epithelium give rise to haploid spermatozoa. During differentiation, spermatozoa move toward the lumen of the tubules as they undergo mitosis, meiosis, and maturation. This process takes 64–74 days, is coordinated by Sertoli cells, and continues throughout life. CLINICAL POINT Semen analysis—used to assess male fertility or successful outcome of vasectomy—measures the amount of semen and other characteristics of sperm cells in an ejaculate. It includes semen volume (commonly 2–6 mL), pH (normally 7.2–7.8), liquefaction time, agglutination (or clumping), sperm cell count (>20 × 106/mL), motility (typically >50% of motile spermatozoa per sample plus an evaluation of forward progression), morphology (size, shape, and appearance of spermatozoa by light microscopy), leukocyte count (normally <1 × 106/mL of ejaculate), and tests for antisperm antibodies and fructose levels (150–600 mg/dL). Sperm culture may also be undertaken to determine signs of inflammation or infection. 17.4 HISTOLOGY OF SEMINIFEROUS TUBULES A distinct connective tissue capsule, a layer of flattened myoid cells, and a basement membrane surround seminiferous tubules. The seminiferous epithelium is an unusual, complex stratified epithelium with two cell populations: spermatogenic (or germ) cells and nonproliferating Sertoli cells. In a seminiferous tubule, germ cells are at various stages of spermatogenesis. The cells closest to the basement membrane with spherical nuclei are spermatogonia. Larger cells with spherical nuclei but with distinctive spaghetti-like chromatin are primary spermatocytes. The haploid secondary spermatocytes are seldom seen; almost as soon as they form they divide and produce spermatids. During a transformation period, spermatids attach to the relatively few Sertoli cells, which are tall and pillar-like. The bases of the Sertoli cells rest on the basement membrane; the free ends of the cells extend radially and reach the lumen. Spermatids, which are known as early and late, do not divide but mature into spermatozoa, which are released into the lumen and carried into efferent ducts. 17.5 ULTRASTRUCTURE OF SEMINIFEROUS TUBULES AND SPERMATOGENESIS Spermatogonia are a continuously renewing stem cell population next to the basement membrane of the seminiferous tubule. They are diploid stem cells, with a diameter of about 12 μm and a relatively large spherical nucleus, and are the most immature group. On division, they give rise to primary spermatocytes, which have relatively large nuclei and are in the middle third of the seminiferous epithelium. After 10–22 days, these cells undergo meiotic division and give rise to smaller secondary spermatocytes, which rapidly undergo a second meiotic division with no DNA replication. The resulting round spermatids have a diameter of about 9 μm and a haploid chromosome number and DNA content. Spermatids are embedded in invaginations of Sertoli cells. As spermatids move toward the tubule lumen, they elongate and undergo an elaborate process of maturation without mitosis, known as spermiogenesis. Resulting spermatozoa are highly specialized cells with a single flagellum and a small, condensed, conical nucleus. Spermatozoa, about 300 million being produced daily, are released into the lumen. CLINICAL POINT Spermatocytic seminoma—an uncommon testicular neoplasm derived from germ cells but clinically and pathologically distinct from other more classic testicular tumors—usually affects men older than 60 years of age and is rarely malignant. Immunocytochemistry of biopsy samples shows positive staining for markers specific for cells undergoing meiosis in addition to genomic abnormalities on chromosome 9, suggesting that primary spermatocytes are cells of origin of this tumor. Orchiectomy is the usual mode of treatment. 17.6 ULTRASTRUCTURE OF GERM CELLS AND EARLY SPERMIOGENESIS Spermatids undergo an elaborate process of maturation known as spermiogenesis. Sequential changes take place whereby spherical, nonmotile spermatids become elongated, motile spermatozoa. Occurring in the upper layers of seminiferous epithelium, these changes include condensation of nuclear chromatin, elongation of the nucleus, formation of the acrosome, migration of cytoplasmic organelles to positions typical of mature cells, formation of a single flagellum, and loss of residual cytoplasm. At first, several small acrosomal vesicles form from the juxtanuclear Golgi complex. They coalesce into a single large membrane-bound acrosome, which adheres to the nuclear envelope. An electron-dense acrosomal granule forms within the vesicle, which gradually spreads to cap the anterior surface of the nucleus and ultimately becomes the front of the mature spermatozoon. The acrosome, a modified lysosome, contains hyaluronidase, lysosomal hydrolases, and protease enzymes that allow spermatozoa to penetrate the corona radiata and zona pellucida of the oocyte in the female at fertilization. CLINICAL POINT Seminomas Only gold members can continue reading. Log In or Register to continue Share this: Share on X (Opens in new window) X Share on Facebook (Opens in new window) Facebook Like this:Like Loading… Related Related posts: CARDIOVASCULAR SYSTEM RESPIRATORY SYSTEM SPECIAL SENSES FEMALE REPRODUCTIVE SYSTEM Stay updated, free articles. Join our Telegram channel Join Tags: Netters Essential Histology Jun 18, 2016 | Posted by admin in HISTOLOGY | Comments Off on MALE REPRODUCTIVE SYSTEM Full access? Get Clinical Tree
17 MALE REPRODUCTIVE SYSTEM 17.1. Overview 17.2. Anatomy and Histology of Testes 17.3. Testicular Development and Spermatogenesis 17.4. Histology of Seminiferous Tubules 17.5. Ultrastructure of Seminiferous Tubules and Spermatogenesis 17.6. Ultrastructure of Germ Cells and Early Spermiogenesis 17.7. Ultrastructure of Germ Cells and Later Spermiogenesis 17.8. Ultrastructure and Function of Sertoli Cells 17.9. Histology and Ultrastructure of Leydig Cells 17.10. Ultrastructure and Function of Leydig Cells 17.11. Anatomy and Histology of the Epididymis 17.12. Histology and Function of the Epididymis 17.13. Histology of the Ductus (Vas) Deferens 17.14. Ultrastructure and Function of the Ductus (Vas) Deferens 17.15. Anatomy and Histology of the Prostate and Seminal Vesicles 17.16. Histology and Function of the Prostate 17.17. Ultrastructure of the Prostate 17.18. Histology of Seminal Vesicles 17.19. Anatomy and Histology of the Urethra and Penis 17.20. Histology of the Penis 17.21. Histology and Function of the Penis 17.1 OVERVIEW The male reproductive system includes the paired primary sex organs, the testes, which have both exocrine and endocrine functions, and several secondary sex organs consisting of excretory ducts and accessory glands. The scrotum and penis, an erectile organ through which the distal urethra passes, are external genitalia. The testes reside outside the body cavity in the scrotum, where they are suspended and maintained in position. Testes and associated spermatic cords are invested by distinct layers of tissue acquired during descent of male gonads from their original retroperitoneal position in the abdominal cavity to the scrotum. Testes contain small convoluted seminiferous tubules, whose germinal epithelium produces male germ cells known as spermatozoa, and interstitial connective tissue. Leydig (interstitial) cells produce testosterone, the hormone responsible for male secondary sex characteristics. After spermatozoa are produced in the testes, they travel a long, tortuous route: from seminiferous tubules to the paired rete testis, efferent ductules (ductuli efferenti), epididymis, ductus (vas) deferens, and ejaculatory ducts, and to the single urethra and penis. The accessory glands include two seminal vesicles, single prostate gland, and paired bulbourethral glands that secrete into ejaculatory ducts and urethra. Three key functions of this system are production of spermatozoa, delivery of these cells via semen into the female reproductive tract, and production of testosterone. CLINICAL POINT The most common causes of male infertility—failure to initiate a pregnancy in a fertile female partner—are a low sperm count, production of immobile or misshapen spermatozoa, and blockages along the male reproductive tract that prevent normal delivery of semen. Other causes include testicular damage caused by cryptorchism (undescended testes), orchitis (inflammation of testes), and varicocele (scrotal varicose veins). Radiation, excessive tobacco use or alcohol consumption, and hormonal disturbances may also contribute to it. 17.2 ANATOMY AND HISTOLOGY OF TESTES The testis, an ovoid gland weighing about 15 g, is encased in a thick capsule of dense fibroelastic connective tissue known as the tunica albuginea, because it appears white in life. An outer visceral layer of the tunica vaginalis invests the capsule externally. Along the posterior border of the testis, the capsule projects inward as a thickened ridge, known as the mediastinum testis. This ridge corresponds to the hilum of other glands—the site where ducts, blood vessels, lymphatics, and nerves connect to the gland interior. Thin fibrous partitions, or septa, radiate from the mediastinum and form wedge-shaped lobules, about 250 in the human. The lobules contain seminiferous tubules, which are sectioned in different planes because they have a convoluted course. Each testis has 600–1200 seminiferous tubules, with a total length of 280–400 m. In the mediastinum, seminiferous tubules empty into tubuli recti and rete testis, which coalesce to form six to eight efferent ductules. These ducts drain testicular fluid and spermatozoa to the proximal part of the epididymis. The rete testis is a labyrinthine network of collecting chambers of simple cuboidal epithelium. Interstitial connective tissue constitutes 20%-30% of the substance of the gland and consists of vascularized connective tissue with clusters of hormone-producing Leydig cells. CLINICAL POINT Orchitis—inflammation of one or both testes—is usually caused by a viral, bacterial, or fungal infection. Viral orchitis, most commonly caused by the mumps virus, typically affects boys after puberty. Most cases of bacterial orchitis occur in young men via sexually transmitted pathogens (e.g., gonorrhea, trachomatis, Chlamydia) or in older men with benign prostatic hypertrophy. Orchitis may also accompany infections spread from other regions (e.g., epididymitis, prostatitis, cystitis). The diagnosis is via urinalysis, urine culture, complete blood count (CBC), and testicular ultrasonography. 17.3 TESTICULAR DEVELOPMENT AND SPERMATOGENESIS Testes derive from embryonic intermediate mesoderm that initially gives rise to primary epithelial sex cords, the precursors of seminiferous tubules. In the 4-week embryo, primordial germ cells migrate from yolk sac endoderm to the cords. Newborn testis consists of solid cords of germ cells arranged in layers and closely associated with epithelial cells that will be supporting Sertoli cells. The cords remain solid until puberty, when they lengthen, increase in diameter, and acquire a lumen. Leydig cells develop from mesenchyme between the seminiferous tubules. At puberty, primitive germ cells—spermatogonia—enlarge and become mitotically active. These cells undergo the process of spermatogenesis, in which diploid spermatogonia in seminiferous epithelium give rise to haploid spermatozoa. During differentiation, spermatozoa move toward the lumen of the tubules as they undergo mitosis, meiosis, and maturation. This process takes 64–74 days, is coordinated by Sertoli cells, and continues throughout life. CLINICAL POINT Semen analysis—used to assess male fertility or successful outcome of vasectomy—measures the amount of semen and other characteristics of sperm cells in an ejaculate. It includes semen volume (commonly 2–6 mL), pH (normally 7.2–7.8), liquefaction time, agglutination (or clumping), sperm cell count (>20 × 106/mL), motility (typically >50% of motile spermatozoa per sample plus an evaluation of forward progression), morphology (size, shape, and appearance of spermatozoa by light microscopy), leukocyte count (normally <1 × 106/mL of ejaculate), and tests for antisperm antibodies and fructose levels (150–600 mg/dL). Sperm culture may also be undertaken to determine signs of inflammation or infection. 17.4 HISTOLOGY OF SEMINIFEROUS TUBULES A distinct connective tissue capsule, a layer of flattened myoid cells, and a basement membrane surround seminiferous tubules. The seminiferous epithelium is an unusual, complex stratified epithelium with two cell populations: spermatogenic (or germ) cells and nonproliferating Sertoli cells. In a seminiferous tubule, germ cells are at various stages of spermatogenesis. The cells closest to the basement membrane with spherical nuclei are spermatogonia. Larger cells with spherical nuclei but with distinctive spaghetti-like chromatin are primary spermatocytes. The haploid secondary spermatocytes are seldom seen; almost as soon as they form they divide and produce spermatids. During a transformation period, spermatids attach to the relatively few Sertoli cells, which are tall and pillar-like. The bases of the Sertoli cells rest on the basement membrane; the free ends of the cells extend radially and reach the lumen. Spermatids, which are known as early and late, do not divide but mature into spermatozoa, which are released into the lumen and carried into efferent ducts. 17.5 ULTRASTRUCTURE OF SEMINIFEROUS TUBULES AND SPERMATOGENESIS Spermatogonia are a continuously renewing stem cell population next to the basement membrane of the seminiferous tubule. They are diploid stem cells, with a diameter of about 12 μm and a relatively large spherical nucleus, and are the most immature group. On division, they give rise to primary spermatocytes, which have relatively large nuclei and are in the middle third of the seminiferous epithelium. After 10–22 days, these cells undergo meiotic division and give rise to smaller secondary spermatocytes, which rapidly undergo a second meiotic division with no DNA replication. The resulting round spermatids have a diameter of about 9 μm and a haploid chromosome number and DNA content. Spermatids are embedded in invaginations of Sertoli cells. As spermatids move toward the tubule lumen, they elongate and undergo an elaborate process of maturation without mitosis, known as spermiogenesis. Resulting spermatozoa are highly specialized cells with a single flagellum and a small, condensed, conical nucleus. Spermatozoa, about 300 million being produced daily, are released into the lumen. CLINICAL POINT Spermatocytic seminoma—an uncommon testicular neoplasm derived from germ cells but clinically and pathologically distinct from other more classic testicular tumors—usually affects men older than 60 years of age and is rarely malignant. Immunocytochemistry of biopsy samples shows positive staining for markers specific for cells undergoing meiosis in addition to genomic abnormalities on chromosome 9, suggesting that primary spermatocytes are cells of origin of this tumor. Orchiectomy is the usual mode of treatment. 17.6 ULTRASTRUCTURE OF GERM CELLS AND EARLY SPERMIOGENESIS Spermatids undergo an elaborate process of maturation known as spermiogenesis. Sequential changes take place whereby spherical, nonmotile spermatids become elongated, motile spermatozoa. Occurring in the upper layers of seminiferous epithelium, these changes include condensation of nuclear chromatin, elongation of the nucleus, formation of the acrosome, migration of cytoplasmic organelles to positions typical of mature cells, formation of a single flagellum, and loss of residual cytoplasm. At first, several small acrosomal vesicles form from the juxtanuclear Golgi complex. They coalesce into a single large membrane-bound acrosome, which adheres to the nuclear envelope. An electron-dense acrosomal granule forms within the vesicle, which gradually spreads to cap the anterior surface of the nucleus and ultimately becomes the front of the mature spermatozoon. The acrosome, a modified lysosome, contains hyaluronidase, lysosomal hydrolases, and protease enzymes that allow spermatozoa to penetrate the corona radiata and zona pellucida of the oocyte in the female at fertilization. CLINICAL POINT Seminomas Only gold members can continue reading. Log In or Register to continue Share this: Share on X (Opens in new window) X Share on Facebook (Opens in new window) Facebook Like this:Like Loading… Related Related posts: CARDIOVASCULAR SYSTEM RESPIRATORY SYSTEM SPECIAL SENSES FEMALE REPRODUCTIVE SYSTEM Stay updated, free articles. Join our Telegram channel Join Tags: Netters Essential Histology Jun 18, 2016 | Posted by admin in HISTOLOGY | Comments Off on MALE REPRODUCTIVE SYSTEM Full access? Get Clinical Tree