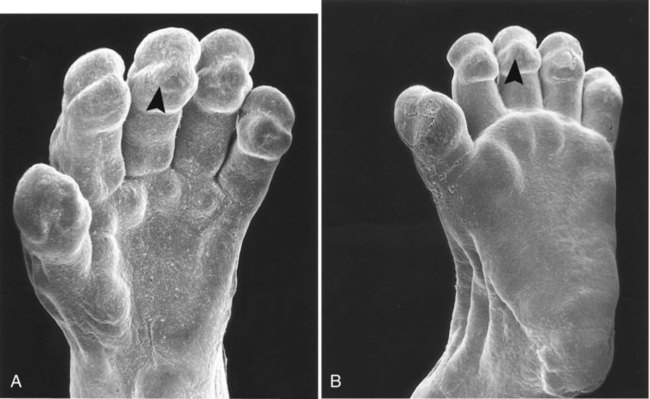
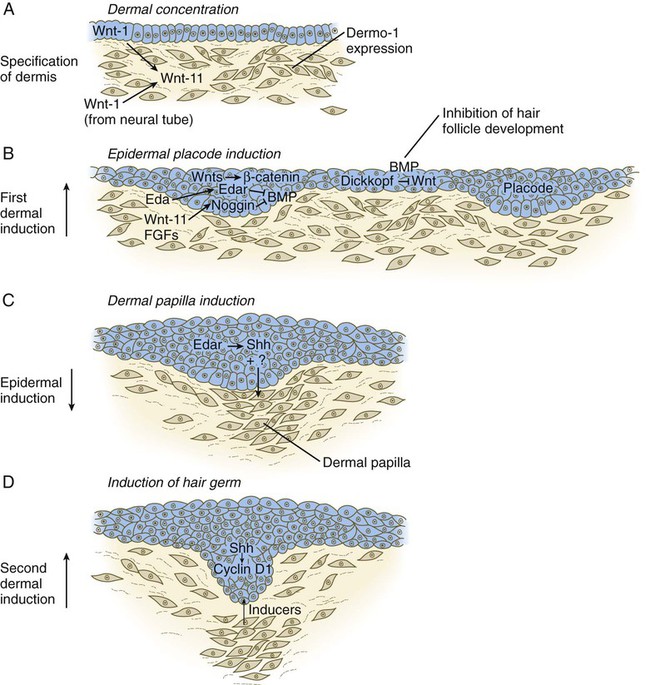
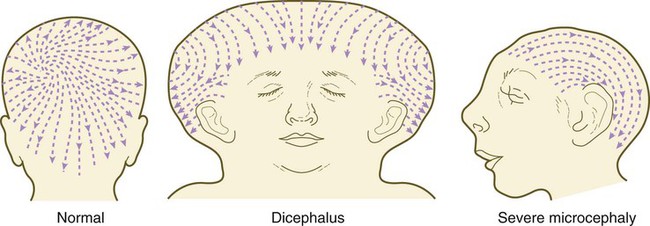
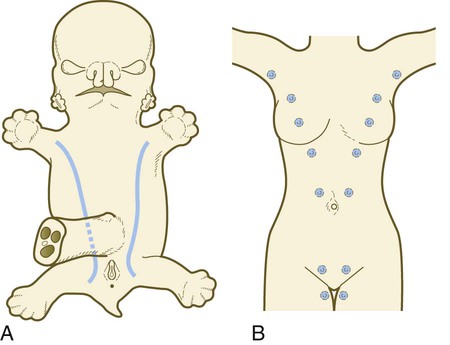
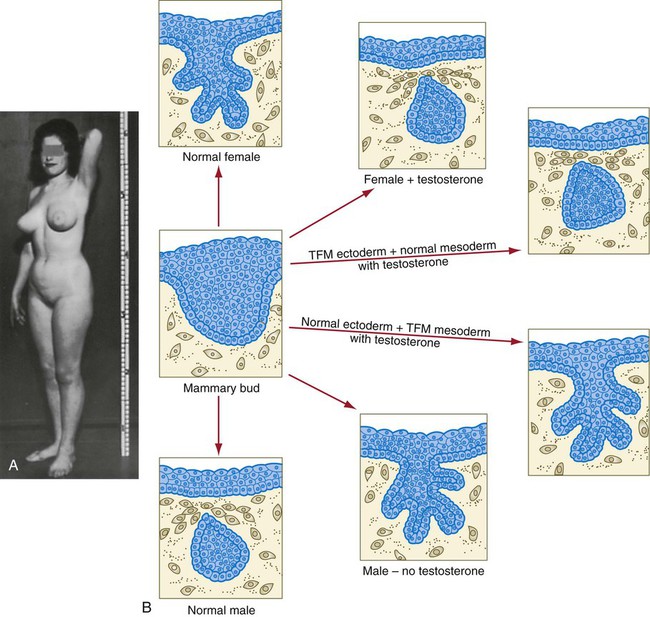
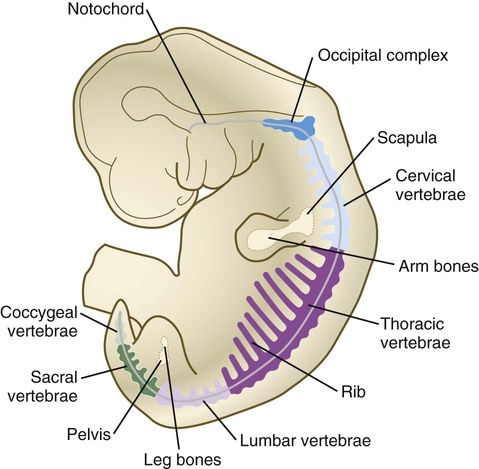
Chapter 9 The construction of the tissues of the body involves developmental phenomena at two levels of organization. One is the level of individual cells, in which the cells that make up a tissue undergo increasing specialization through a process called cytodifferentiation (see discussion of restriction, determination, and differentiation, [p. 85]). At the next level of complexity, various cell types develop in concert to form specific tissues through a process called histogenesis. This chapter discusses the development of three important tissues of the body: skin, bone, and muscle. The histogenesis of each of these tissues exemplifies important aspects of development. The skin, consisting of the epidermis and dermis, is one of the largest structures in the body. The epidermis represents the interface between the body and its external environment, and its structure is well adapted for local functional requirements. Simple inspection of areas such as the scalp and palms shows that the structure of the integument varies from one part of the body to another. These local variations result from inductive interactions between the ectoderm and underlying mesenchyme. Abnormalities associated with the integumentary system are presented later in Clinical Correlation 9.1. Clinical Correlation 9.1 Abnormalities of Skin Development Several types of anomalies affect the integumentary system. Ectodermal dysplasia is a germ layer defect that can affect many ectodermal derivatives depending on the type and severity of the condition. In addition to abnormalities of the epidermis itself, this syndrome can include the absence or abnormalities of hair and teeth (see Fig. 8.19) and short stature (caused by anterior pituitary gland maldevelopment). Several specific defects in the synthesis of types I and III collagen are lumped together as variants of the Ehlers-Danlos syndrome. Individuals with this condition typically have hypermobile joints. Affected skin can be characterized by sagging or hyperelasticity (Fig. 9.15). Angiomas of the skin (birthmarks) are vascular malformations characterized by localized red or purplish spots ranging in size from tiny dots to formations many inches in diameter. Angiomas consist of abnormally prominent plexuses of blood vessels in the dermis, and they may be raised above the level of the skin or a mucous membrane (see Fig. 17-49). The outer layer of the skin begins as a single layer of ectodermal cells (Fig. 9.1A). As development progresses, the ectoderm becomes multilayered, and regional differences in structure become apparent. The first stage in epidermal layering is the formation of a thin outer layer of flattened cells known as the periderm at the end of the first month of gestation (Fig. 9.1B). Cells of the periderm, which is present in the epidermis of all amniote embryos, seem to be involved in the exchange of water, sodium, and possibly glucose between the amniotic fluid and the epidermis. By the third month, the epidermis becomes a three-layered structure, with a mitotically active basal (or germinative) layer, an intermediate layer of cells (Fig. 9.1D) that represent the progeny of the dividing stem cells of the basal layer, and a superficial layer of peridermal cells bearing characteristic surface blebs (Fig. 9.2). Peridermal cells contain large amounts of glycogen, but the function of this glycogen remains uncertain. When the multilayered epidermis becomes established, a regular cellular organization and sequence of differentiation appear within it (Fig. 9.3). Stem cells* of the basal layer (stratum basale) divide and contribute daughter cells to the next layer, the stratum spinosum. The movement of epidermal cells away from the basal layer is preceded by a loss of adhesiveness to basal lamina components (e.g., fibronectin, laminin, and collagen types I and IV). These cellular properties can be explained by the loss of several integrins, which attach the basal cells to the underlying basal lamina. Cells of the stratum spinosum produce prominent bundles of keratin filaments, which converge on the patchlike desmosomes binding the cells to each other. The formation of epidermal ridges is closely associated with the earlier appearance of volar pads on the ventral surfaces of the fingers and toes (Fig. 9.4). Volar pads first form on the palms at about When the epidermal ridges first form, the tips of the digits are still smooth, and the fetal epidermis is covered with peridermal cells. Beneath the smooth surface, however, epidermal and dermal ridges begin to take shape (Fig. 9.5). Late in the fifth month of pregnancy, the epidermal ridges become recognizable features of the surface landscape. The dermis arises from several sources. In the trunk, dorsal dermis arises from the dermatome of the somites, whereas ventral and lateral dermis and dermis of the limbs is derived from the lateral plate mesoderm. In the face, much of the cranial skin, and anterior neck, dermal cells are descendants of cranial neural crest ectoderm (see Fig. 12.9). Ectodermal Wnt signaling, acting through the β-catenin pathway, specifies the dermomyotomal cells, as well as mesenchymal cells of the ventral somatopleure closest to the ectoderm, to become dermal cells, which express the dermal marker, Dermo 1 (see Fig. 9.8A). The future dermis is initially represented by loosely aggregated mesenchymal cells that are highly interconnected by focal tight junctions on their cellular processes. These early dermal precursors secrete a watery intercellular matrix rich in glycogen and hyaluronic acid. If ectoderm from one part of the body is combined with dermis from another area, the ectoderm differentiates into a regional pattern characteristic of underlying dermis, rather than a pattern appropriate for the site of origin of the ectoderm (Fig. 9.6). Cross-species recombination experiments have shown that, even in distantly related animals, skin ectoderm and mesenchyme can respond to each other’s inductive signals. As a result of inductive influences by the dermis, the epidermis produces a wide variety of appendages, such as hair, nails, sweat and sebaceous glands, mammary glands, and the enamel component of teeth. (The development of teeth is discussed in Chapter 14.) Hair formation is first recognizable at about the twelfth week of pregnancy as regularly spaced epidermal placodes associated with small condensations of dermal cells called dermal papillae (Fig. 9.7). Under the continuing influence of a dermal papilla, the placode forms an epidermal downgrowth (hair germ), which over the next few weeks forms an early hair peg. In succeeding weeks, the epidermal peg overgrows the dermal papilla, and this process results in the shaping of an early hair follicle. At this stage, the hair follicle still does not protrude beyond the outer surface of the epidermis, but in the portion of the follicle that penetrates deeply into the dermis, two bulges presage the formation of sebaceous glands, which secrete an oily skin lubricant (sebum), and are the attachment site for the tiny arrector pili muscle. The arrector pili is a mesodermally derived smooth muscle that lifts the hair to a nearly vertical position in a cold environment. In many animals, this increases the insulation properties of the hair. The developing hair follicle induces the adjacent dermal mesoderm to form the smooth muscle cells of this muscle. As the developing hair matures, a small bulge below the sebaceous gland marks an aggregation of epidermal stem cells (Fig. 9.7D). The formation of a hair involves a series of inductive interactions mediated by signals that are only partly understood. When a dense condensation of dermal cells has formed beneath the ectoderm (Fig. 9.8A), the first of two dermal inductions results in the thickening of ectoderm in very regularly arranged locations to form epidermal placodes (Fig. 9.8B). Fibroblast growth factor (FGF) and Wnt (mainly Wnt-11) signaling from the dermis, along with the inactivation of local bone morphogenetic proteins (BMPs), stimulates the activation of other Wnts in the ectoderm to form an epidermal placode. The response of the ectoderm is to produce other Wnts, acting through β-catenin intermediates, and Edar, the receptor for the signaling molecule ectodysplasin. In the areas where hairs will not develop (interfollicular areas), placode formation is inhibited by locally produced BMPs and by the inhibition of Wnts by Dickkopf. How the epidermal placodes are spaced in such a geometrically regular fashion is still not well understood. The newly formed epidermal placodes become the inducing agent and stimulate the aggregation of mesenchymal cells beneath the placode to form the dermal papilla (Fig. 9.8C). Sonic hedgehog, produced by the epidermal placode, seems to be involved in this induction, but the identity of other signals is unknown. Next, the dermal papilla initiates the second dermal induction by stimulating downgrowth of the cells of the epidermal placode into the dermis (Fig. 9.8D). Epidermal downgrowth, which involves considerable epidermal cellular proliferation, is stimulated by expression of sonic hedgehog by the epidermal cells and the subsequent expression of cyclin D1, part of the cell cycling pathway. Later formation of a hair is structurally and biochemically an extremely complex process, which, among other things, involves the expression of a range of Hox genes in specific locations and at specific times along the length of each developing hair. Once formed, an individual hair follows a regular cycle of growth and shedding (see Fig. 9.7). During anagen, the first phase in the cycle, the hair is actively growing (around 10 cm per year). This phase can last up to 5 to 6 years. Then it enters catagen, a phase lasting 1 or 2 weeks, during which the hair follicle regresses to only a fraction of its original length. The hair stops growing in the resting phase (telogen), which lasts 5 to 6 weeks, after which time the hair is shed (exogen). Adjacent hairs are frequently in different phases of the hair cycle. The pattern of epidermal appendages such as hairs has been shown experimentally to relate to patterns generated in the dermis. Other studies have compared patterns of scalp hairs between normal embryos and embryos with cranial malformations (Fig. 9.9) and have shown a correlation between whorls and the direction of hair growth and the tension on the epidermis at the time of formation of the hair follicles. Toward the end of the third month, epidermal thickenings (primary nail field) on the dorsal surfaces of the digits mark the beginnings of nail development. Cells from the primary nail field expand proximally to undercut the adjacent epidermis (Fig. 9.10). Proliferation of cells in the proximal part of the nail field results in the formation of a proximal matrix, which gives rise to the nail plate that grows distally to cover the nail bed. The nail plate itself consists of highly keratinized epidermal cells. A thin epidermal layer, the eponychium, initially covers the entire nail plate, but it eventually degenerates, except for a thin persisting rim along the proximal end of the nail. The thickened epidermis underlying the distalmost part of the nail is called the hyponychium, and it marks the border between dorsal and ventral skin. Outgrowing fingernails reach the ends of the digits by about 32 weeks, whereas in toenails, this does not occur until 36 weeks. As with many glandular structures, the mammary glands arise as epithelial (in this case, ectodermal) downgrowths into mesenchyme in response to inductive influences by the mesenchyme. The first morphological evidence of mammary gland development is the appearance of two bands of ectodermal thickenings called milk lines (part of the wolffian ridge [see p. 111]) running along the ventrolateral body walls in embryos of both genders at about 6 weeks (Fig. 9.11A). They are marked by the expression of various Wnts within the ectodermal cells. The thickened ectoderm of the milk lines undergoes fragmentation, and remaining areas form the primordia of the mammary glands. The craniocaudal level and the extent along the milk lines at which mammary tissue develops vary among species. Comparing the location of mammary tissue in cows (caudal), humans (in the pectoral region), and dogs (along the length of the milk line) shows the wide variation in location and number of mammary glands. In humans, supernumerary mammary tissue or nipples can be found anywhere along the length of the original milk lines (Fig. 9.11B). Individual mammary placodes form from aggregation and proliferation of ectodermal cells of the milk line under the inductive influence of the signaling molecule neuregulin-3. Their dorsoventral location is marked by the expression of the transcription factor Tbx-3. Mammary ductal epithelial downgrowths (Fig. 9.12) are associated with two types of mesoderm: fibroblastic and fatty. The early epithelial downgrowth secretes parathyroid hormone–related hormone, which increases the sensitivity of the underlying mesenchymal cells to BMP-4. BMP-4 signals within the underlying mesenchyme have two principal effects (see Fig. 9.12B). First, they stimulate further downgrowth of the mammary epithelial bud. Second, they stimulate the expression of the transcription factor Msx-2, which inhibits the formation of hair follicles in the region of the nipple. Experimental evidence suggests that inductive interactions with the fatty component of the connective tissue are responsible for the characteristic shaping of the mammary duct system. As with many developing glandular structures, the inductive message seems to be mediated to a great extent by the extracellular matrix of the connective tissue. The role of the mesoderm and testosterone receptors is well illustrated in experiments involving mice with a genetic mutant, androgen insensitivity syndrome. This is the counterpart of a human condition called the testicular feminization syndrome, in which genetic male individuals lack testosterone receptors. Despite having high circulating levels of testosterone, these individuals develop female phenotypes, including typical female breast development (Fig. 9.13A), because without receptors, the tissue cannot respond to the testosterone. In vitro recombination experiments on mice with androgen insensitivity have been instrumental in understanding the role of the mesoderm in mediating the effects of testosterone on mammary duct development (Fig. 9.13B). If mutant mammary ectoderm is combined with normal mesoderm in the presence of testosterone, the mammary ducts regress, but normal ectoderm combined with mutant mesoderm continues to form normal mammary ducts despite being exposed to high levels of testosterone. This shows that the genetic defect in testicular feminization is expressed in the mesoderm. The postnatal development of female mammary gland tissue is also highly responsive to its hormonal environment. The simple mammary duct system that was laid down in the embryo remains in an infantile condition until it is exposed to the changing hormonal environment at the onset of puberty (Fig. 9.14A). Increasing levels of circulating estrogens, acting on a base of growth hormone and insulinlike growth factor activity, stimulate the proliferation of the mammary ducts and enlargement of the pad of fatty tissue that underlie it (Fig. 9.14B). As is the case with testosterone effects, estrogen effects on the epithelium of the mammary ducts are mediated via paracrine influences from the mammary connective tissue stroma, which contains the estrogen receptors. The next major change in the complete cycle of mammary tissue development occurs during pregnancy, although minor cyclical changes in mammary tissue are detectable in each menstrual cycle. During pregnancy, increased amounts of progesterone, along with prolactin and placental lactogen, stimulate the development of secretory alveoli at the ends of the branched ducts (Fig. 9.14C). With continuing development of the alveoli, the epithelial cells build up increased numbers of the cytoplasmic organelles, such as rough endoplasmic reticulum and the Golgi apparatus, which are involved in protein synthesis and secretion. Lactation involves numerous reciprocal influences between the mammary glands and the brain; these are summarized in Figure 9.14D. Stimulated by prolactin secretion from the anterior pituitary, the alveolar cells synthesize milk proteins (casein and α-lactalbumin) and lipids. In a rapid response to the suckling stimulus, the ejection of milk is triggered by the release of oxytocin by the posterior portion of the pituitary. Oxytocin causes the contraction of myoepithelial cells, which surround the alveoli. Suckling also causes an inhibition of the release of luteinizing hormone–releasing hormone by the hypothalamus that results in the inhibition of ovulation and a natural form of birth control. With cessation of nursing, reduced prolactin secretion and the inhibitory effects of nonejected milk in the mammary alveoli result in the cessation of milk production. The mammary alveoli regress, and the duct system of the mammary gland returns to the nonpregnant state (Fig. 9.14E). Clinical Correlation 9.1 summarizes several types of anomalies that affect the integumentary system. The deep skeletal elements of the body typically first appear as cartilaginous models of the bones that will ultimately be formed (Fig. 9.16). At specific periods during embryogenesis, the cartilage is replaced by true bone through the process of endochondral ossification. In contrast, the superficial bones of the face and skull form by the direct ossification of mesenchymal cells without an intermediate cartilaginous stage (intramembranous bone formation). Microscopic details of intramembranous and endochondral bone formation are presented in standard histology texts and are not repeated here. To differentiate into defined skeletal elements, the mesenchymal precursor cells must often interact with elements of their immediate environment—typically epithelia with associated basal laminae—or components of the neighboring extracellular matrix. Details of the interactions vary among regions of the body. In the limb, a continuous interaction between the apical ectodermal ridge (see Chapter 10) and the underlying limb bud mesoderm is involved in the specification of the limb skeleton. An inductive interaction between the sclerotome and notochord or neural tube initiates skeletogenesis of the vertebral column. In the head, preskeletal cells of the neural crest may receive information at levels ranging from the neural tube itself, to sites along their path of migration, to the region of their final destination. Inductive interactions between regions of the brain and the overlying mesenchyme stimulate formation of the membrane bones of the cranial vault. Regardless of the nature of the initial induction, the formation of skeletal elements begins along a common path, which diverges into osteogenic or chondrogenic programs, depending on the nature of the immediate environment. Shortly after induction, the preskeletal mesenchymal cells produce the cellular adhesion molecule N-cadherin, which promotes their transformation from a mesenchymal to an epitheliumlike morphology and their forming cellular condensations (Fig. 9.17). The growth factor transforming growth factor-β stimulates the synthesis of fibronectin and finally N-CAM, which maintains the aggregated state of the cells in the preskeletal condensation. At this point, specific differentiation programs come into effect. If the skeletal element is destined to form membranous bone, the transcription factor Runx-2 sets off an osteogenic program (see Fig. 9.17). Osterix (Osx) is a downstream transcription factor from Runx-2 and is also required for the differentiation of osteoblasts. The protein encoded by the Runx2 gene has been shown to control the differentiation of mesenchymal cells into osteoblasts (bone-forming cells). These cells produce molecules characteristic of bone (type I collagen, osteocalcin, and osteopontin) and form spicules of intramembranous bone. If the cellular condensation is destined to form cartilage, it follows the chondrogenic program. Under the influence of Sox-9, the chondroblasts begin to form type II collagen and secrete a cartilaginous matrix (see Fig. 9.17). Some embryonic cartilage (e.g., in the nose, ear, and intervertebral surfaces) remains as permanent cartilage and continues to express Sox-9. The cartilage that forms the basis for endochondral bone formation undergoes specific changes that ultimately promote bone formation around it. A first step is hypertrophy, which occurs under the influence of Runx-2 and the signaling factors Indian hedgehog and BMP-6. The formation of type X collagen is characteristic of hypertrophying cartilage. Then the hypertrophic chondrocytes themselves begin to produce bone proteins, such as osteocalcin, osteonectin, and osteopontin. They also express vascular endothelial growth factor, which stimulates the ingrowth of blood vessels into the hypertrophic cartilage. This sets the stage for the replacement of the eroded hypertrophic cartilage by true bone as osteoblasts accompany the invading capillaries. FGF-18, produced by the perichondrium, inhibits the maturation of the chondrocytes around the periphery of the mass of cartilage as those at the center are undergoing hypertrophy. The earliest stages in establishing the axial skeleton are introduced in Chapter 6. Formation of the axial skeleton is more complex, however, than the simple subdivision of the paraxial mesoderm into somites and the medial displacement of sclerotomal cells to form primordia of the vertebrae. Each vertebra has a complex and unique morphology specified by controls operating at several levels and during several developmental periods. According to the traditional view of vertebral development (see Fig. 6.13
Integumentary, Skeletal, and Muscular Systems
Integumentary System
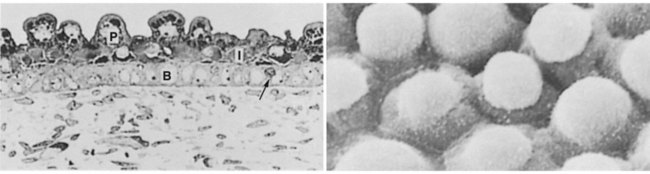
Epidermis
Structural Development
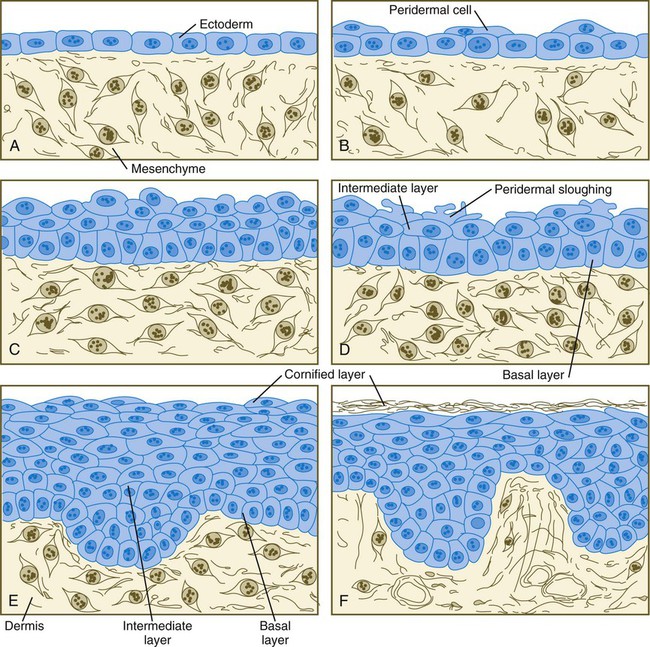
A, At 1 month. B, At 2 months. C, At 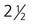 months. D, At 4 months. E, At 6 months. F, After birth. (Adapted from Carlson B: Patten’s foundations of embryology, ed 6, New York, 1996, McGraw-Hill.)
months. D, At 4 months. E, At 6 months. F, After birth. (Adapted from Carlson B: Patten’s foundations of embryology, ed 6, New York, 1996, McGraw-Hill.)
Epidermal Differentiation
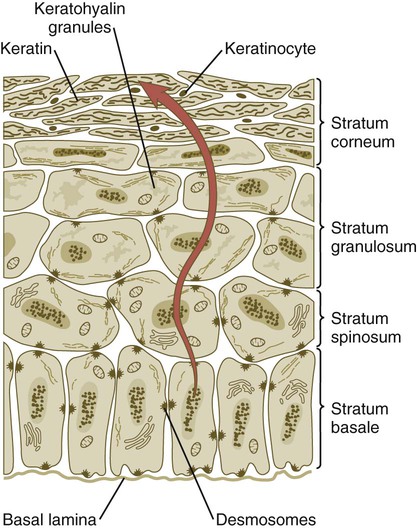
Cells arising in the stratum basale undergo terminal differentiation into keratinocytes as they move toward the surface. (Adapted from Carlson B: Patten’s foundations of embryology, ed 6, New York, 1996, McGraw-Hill.)
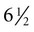 weeks, and by
weeks, and by 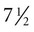 weeks, they have formed on the fingers. The volar pads begin to regress by about
weeks, they have formed on the fingers. The volar pads begin to regress by about 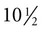 weeks, but while they are present, they set the stage for the formation of the epidermal ridges, which occurs between 11 and 17 weeks. Similar events in the foot occur approximately 1 week later than those in the hand.
weeks, but while they are present, they set the stage for the formation of the epidermal ridges, which occurs between 11 and 17 weeks. Similar events in the foot occur approximately 1 week later than those in the hand.
A, Low-power view of the palmar surface of a digit. B, Epidermal surface of the dermis of the fingertip showing the primary dermal ridges. C, Basal surface of the epidermis showing the epidermal ridges. Bars, 100 µm. (From Misumi Y, Akiyoshi T: Am J Anat 119:419, 1991.)
Dermis
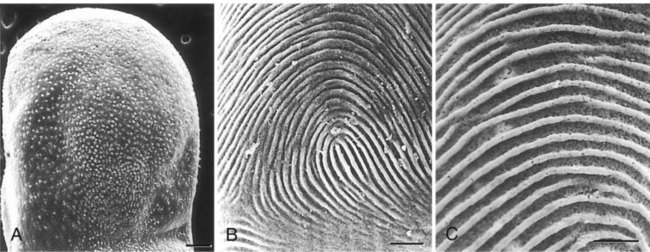
Dermal-Epidermal Interactions
When separated (A), ectoderm and underlying mesenchyme do not differentiate. Recombinations (B and C) show that the dermis determines the nature of the ectodermal differentiation.
Epidermal Appendages
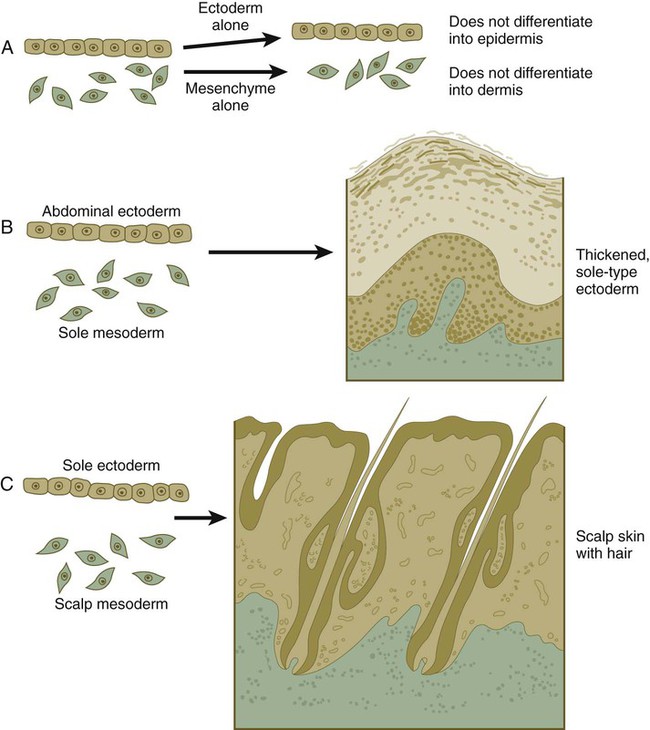
Hair
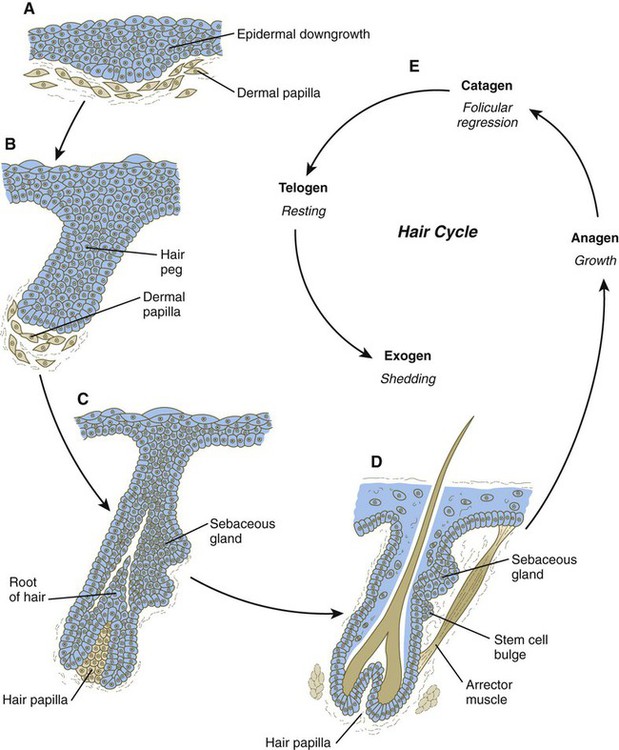
A, Hair primordium (12 weeks). B, Early hair peg (15 to 16 weeks). C, Bulbous hair follicle (18 weeks). D, Adult hair. E, The adult hair cycle.
Nails
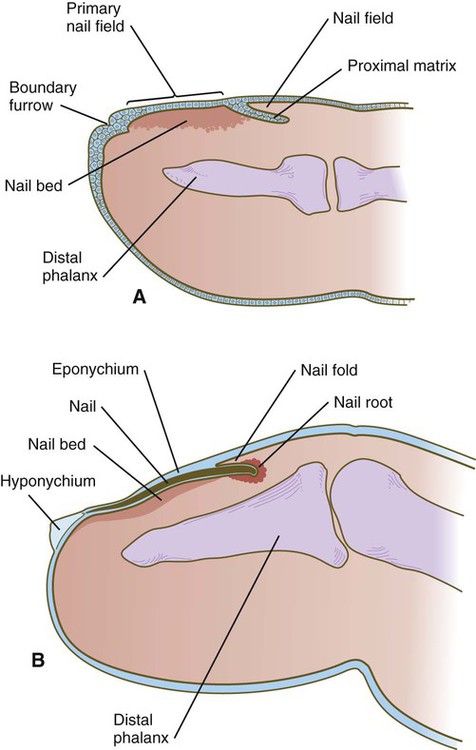
A, In the fourth month, the primary nail field overlies a mesenchymal nail bed and extends proximally as the proximal matrix. B, Close to term, the nail has grown close to the end of the fingertip. Much of the nail is covered by a thin eponychium, most of which will eventually degenerate.
Mammary Glands
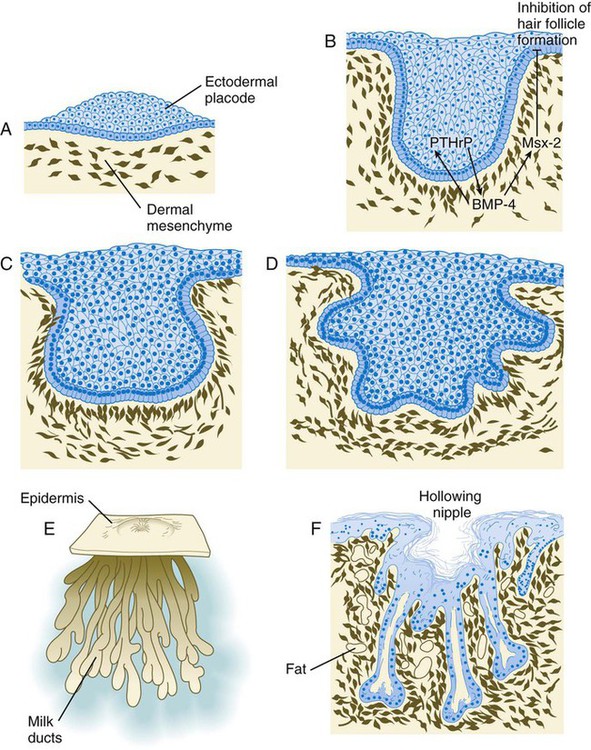
A, Sixth week. B, Seventh week. BMP-4, bone morphogenetic protein-4; PTHrP, parathyroid hormone–related protein. C, Tenth week. D, Fourth month. E, Sixth month. F, Eighth month.
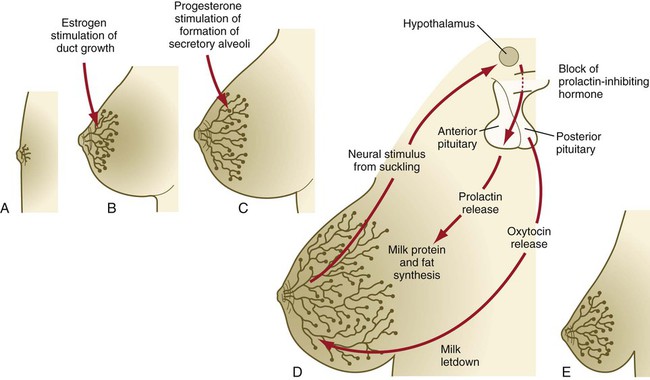
A, Newborn. B, Young adult. C, Adult. D, Lactating adult. E, Adult after lactation.
Skeleton
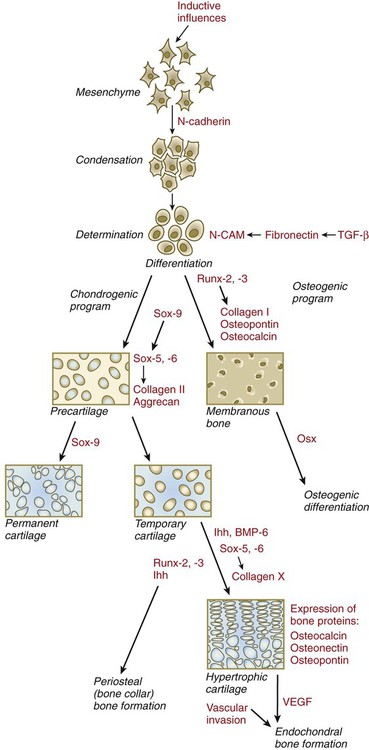
BMP-6, bone morphogenetic protein-6; CAM, cellular adhesion molecule; Ihh, Indian hedgehog; Osx, osterix; TGF-β, transforming growth factor-β; VEGF, vascular endothelial growth factor.
Axial Skeleton
Vertebral Column and Ribs
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Integumentary, Skeletal, and Muscular Systems