57
Immunity
CHAPTER CONTENTS
FUNCTION OF THE IMMUNE SYSTEM
The main function of the immune system is to prevent or limit infections, fungi, and parasites, such as protozoa and worms. The first line of defense against microorganisms is the intact skin and mucous membranes. If microorganisms breach this line and enter the body, then the innate arm of the immune system (second line of defense) is available to destroy the invaders. Because the components of the innate arm (Table 57–1) are preformed and fully active, they can function immediately upon entry of the microorganisms. The ability of the innate arm to kill microorganisms is not specific. For example, a neutrophil can ingest and destroy many different kinds of bacteria.
TABLE 57–1 Main Components of Innate and Adaptive Immunity That Contribute to Humoral (Antibody-Mediated) Immunity and Cell-Mediated Immunity

Highly specific protection is provided by the adaptive (acquired) arm of the immune system (third line of defense), but it takes several days for this arm to become fully functional. The two components of the adaptive arm are cell-mediated immunity and antibody-mediated (humoral) immunity. An overview of the functions and interactions between many of the important members of the innate and adaptive arms of the immune response is provided in Figure 57–1. (The features of the innate and the adaptive arms of the immune system are contrasted in Table 57–2.)

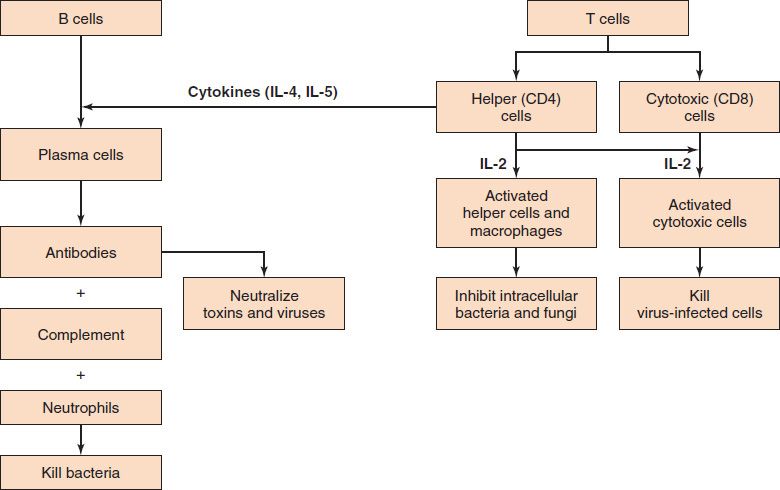
FIGURE 57–1 Introduction to the interactions and functions of the major components of the immune system. Left: Antibody-mediated (humoral) immunity. This is our main defense against extracellular, encapsulated, pyogenic bacteria such as staphylococci and streptococci. Antibodies also neutralize toxins, such as tetanus toxin, as well as viruses, such as hepatitis B virus. Right: Cell-mediated immunity. There are two distinct components. (1) Helper T cells and macrophages are our main defense against intracellular bacteria, such as Mycobacterium tuberculosis, and fungi, such as Histoplasma capsulatum. (2) Cytotoxic T cells are an important defense against viruses and act by destroying virus-infected cells. (IL-4 and IL-5 are interleukin-4 and interleukin-5, respectively.)
The cell-mediated arm consists primarily of T lymphocytes (e.g., helper T cells and cytotoxic T cells), whereas the antibody-mediated arm consists of antibodies (immunoglobulins) and B lymphocytes (and plasma cells). Some of the major functions of T cells and B cells are shown in Table 57–3.
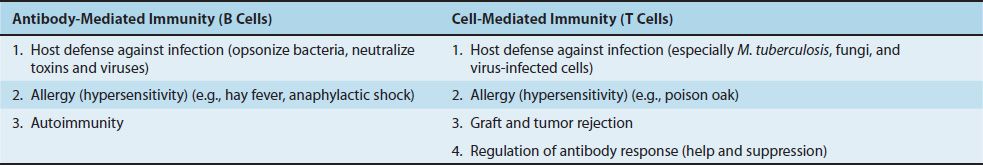
The main functions of antibodies are (1) to neutralize toxins and viruses and (2) to opsonize bacteria, making them easier to phagocytize. Opsonization is the process by which immunoglobulin G (IgG) antibody and the C3b component of complement enhance phagocytosis (see Figure 8–3). Cell-mediated immunity, on the other hand, inhibits organisms such as fungi, parasites, and certain intracellular bacteria such as Mycobacterium tuberculosis; it also kills virus-infected cells and tumor cells.
Both the cell-mediated and antibody-mediated responses are characterized by three important features: (1) they exhibit remarkable diversity (i.e., they can respond to millions of different antigens); (2) they have a long memory (i.e., they can respond many years after the initial exposure because memory T cells and memory B cells are produced); and (3) they exhibit exquisite specificity (i.e., their actions are specifically directed against the antigen that initiated the response).
The combined effects of certain cells (e.g., T cells, B cells, macrophages, and neutrophils) and certain proteins (e.g., interleukins, antibodies, and complement) produce an inflammatory response, one of the body’s main defense mechanisms. The process by which these components interact to cause inflammation is described in Chapter 8.
Macrophages and certain other phagocytic cells such as dendritic cells participate in both the innate and adaptive arms of the immune response. They are, in effect, a bridge between the two arms. As part of the innate arm, they ingest and kill various microbes. They also present antigen to helper T cells, which is the essential first step in the activation of the adaptive arm (see later). It is interesting to note that neutrophils, which are also phagocytes and have excellent microbicidal abilities, do not present antigen to helper T cells and therefore function in innate but not acquired immunity.
SPECIFICITY OF THE IMMUNE RESPONSE
Cell-mediated immunity and antibody are both highly specific for the invading organism. How do these specific protective mechanisms originate? The process by which these host defenses originate can be summarized by three actions: (1) the recognition of the foreign organism by specific immune cells, (2) the activation of these immune cells to produce a specific response (e.g., antibodies), and (3) the response that specifically targets the organism for destruction. The following examples briefly describe how specific immunity to microorganisms occurs. An overview of these processes with a viral infection as the model is shown in Figure 57–2. A detailed description is presented in Chapter 58.
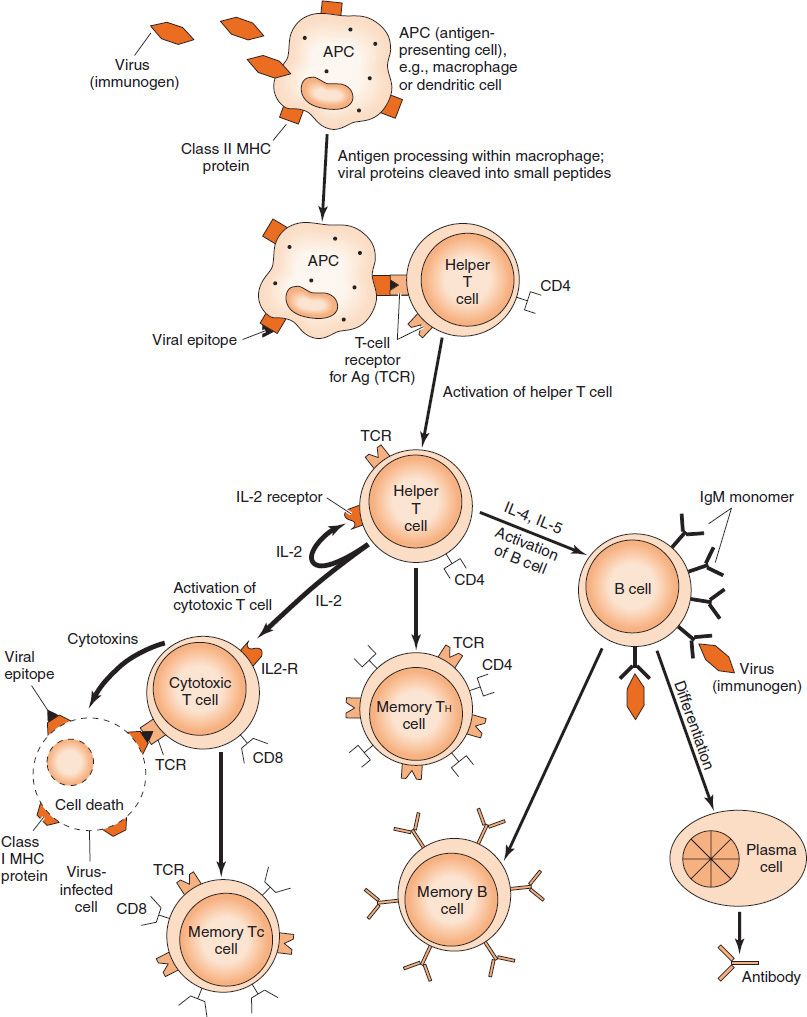
FIGURE 57–2 Overview of the process by which cell-mediated immunity and antibody-mediated immunity are induced by exposure to a virus. Note that the figure shows a virus as the immunogen in the top left corner, but the same processes occur for other microbes, such as bacteria or fungi. IL, interleukin; MHC, major histocompatibility complex. (Modified and reproduced with permission from Stites D, Terr A, Parslow T, eds. Basic & Clinical Immunology. 9th ed. Originally published by Appleton & Lange. Copyright 1997 McGraw-Hill.)
1. Cell-Mediated Immunity
In the following example, a bacterium (e.g., Mycobacterium tuberculosis) enters the body and is ingested by a macrophage. The bacterium is broken down, and fragments of it called antigens or epitopes appear on the surface of the macrophage in association with class II major histocompatibility complex (MHC) proteins. The antigen–class II MHC protein complex interacts with an antigen-specific receptor on the surface of a helper T lymphocyte. Activation and clonal proliferation of this antigen-specific helper T cell occur as a result of the production of interleukins, the most important of which are interleukin-2 (T cell growth factor) and gamma interferon (activates macrophages). These activated helper T cells, aided by activated macrophages, mediate one important component of cellular immunity (i.e., a delayed hypersensitivity reaction specifically against M. tuberculosis).
Cytotoxic (cytolytic) T lymphocytes are also specific effectors of the cellular immune response, particularly against virus-infected cells. In this example, a virus (e.g., influenza virus) is inhaled and infects a cell of the respiratory tract. Viral envelope glycoproteins appear on the surface of the infected cell in association with class I MHC proteins. A cytotoxic T cell binds via its antigen-specific receptor to the viral antigen–class I MHC protein complex and is stimulated to grow into a clone of cells by interleukin-2 produced by helper T cells. These cytotoxic T cells specifically kill influenza virus–infected cells (and not cells infected by other viruses) by recognizing viral antigen–class I MHC protein complexes on the cell surface and releasing perforins that destroy the membrane of the infected cell.
2. Antibody-Mediated Immunity
Antibody synthesis typically involves the cooperation of three cells: antigen-presenting cells (e.g., dendritic cells and macrophages), helper T cells, and B cells. After processing by an antigen-presenting cell, fragments of antigen appear on the surface of that cell in association with class II MHC proteins. The antigen–class II MHC protein complex binds to receptors on the surface of a helper T cell specific for that antigen. This activates the helper T cells to produce interleukins such as interleukin-2 (IL-2), IL-4, and IL-5. These interleukins activate the B cell to produce antibodies specific for that antigen. (Note that the interleukins are nonspecific; the specificity lies in the T cells and B cells and is mediated by the antigen receptors on the surface of these cells.) The activated B cell proliferates and differentiates to form many plasma cells that secrete large amounts of immunoglobulins (antibodies).
Although antibody formation usually involves helper T cells, certain antigens (e.g., bacterial polysaccharides) can activate B cells directly, without the help of T cells, and are called T-cell–independent antigens. In this T-cell–independent response, only IgM is produced by B cells because it requires IL-4 and IL-5 made by the helper T cell for the B cell to “class switch” to produce IgG, IgA, and IgE. See Chapter 59 for a discussion of “class switching,” the process by which the B cell switches the antibody it produces from IgM to one of the other classes.
Figure 57–3 summarizes the human host defenses against virus-infected cells and illustrates the close interaction of various cells in mounting a coordinated attack against the pathogen. The specificity of the response is provided by the antigen receptor (T-cell receptor [TCR]) on the surface of both the CD4-positive T cell and the CD8-positive T cell and by the antigen receptor (IgM) on the surface of the B cell. The interleukins, on the other hand, are not specific.
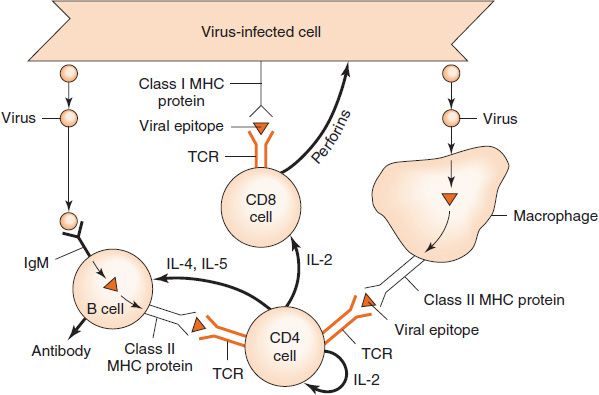
FIGURE 57–3 Induction of cell-mediated immunity and antibody against a viral infection. Right: Virus released by an infected cell is ingested and processed by an antigen-presenting cell (APC) (e.g., a macrophage). The viral epitope is presented in association with a class II major histocompatibility complex (MHC) protein to the virus-specific T-cell receptor (TCR) on the CD4 cell. The macrophage makes interleukin (IL) -1, which helps activate the CD4 cell. The activated CD4 cell makes interleukins (e.g., IL-2, which activates the CD8 cell to attack the virus-infected cell, and IL-4 and IL-5, which activate the B cell to produce antibody). The specificity of the cytotoxic response mounted by the CD8 cell is provided by its TCR, which recognizes the viral epitope presented by the virus-infected cell in association with a class I MHC protein. Left: Virus released by an infected cell interacts with the antigen receptor (IgM monomer) specific for that virus located on the surface of a B cell. The virus is internalized, and the viral proteins are broken down into small peptides. B cells (as well as macrophages) can present viral epitopes in association with class II MHC proteins and activate CD4 cells. The CD4-positive helper cell produces IL-4 and IL-5, which induce the B cell to differentiate into a plasma cell that produces antibody specifically against this virus.
As depicted in Figure 57–3, B cells can perform two important functions during the induction process: (1) they recognize antigens with their surface IgM that acts as an antigen receptor, and (2) they present epitopes to helper T cells in association with class II MHC proteins. Note that the IgM antigen receptor on the B cell can recognize not only foreign proteins but also carbohydrates, lipids, DNA, RNA, and other types of molecules. The class II MHC proteins of the B cell, however, can only present peptide fragments to the helper T cells. This distinction will become important when haptens are discussed later in this chapter. It is this remarkable ability of the IgM antigen receptor on the B cell to bind to an incredibly broad range of molecules that enables B cells to produce antibodies against virtually every molecule known. How the B cell generates such a diverse array of antibodies is described on page 512.
INNATE & ADAPTIVE IMMUNITY
Our immune host defenses can be divided into two major categories: innate (natural) and adaptive (acquired). The features of these two important components of our host defenses are compared in Table 57–2.
1. Innate Immunity
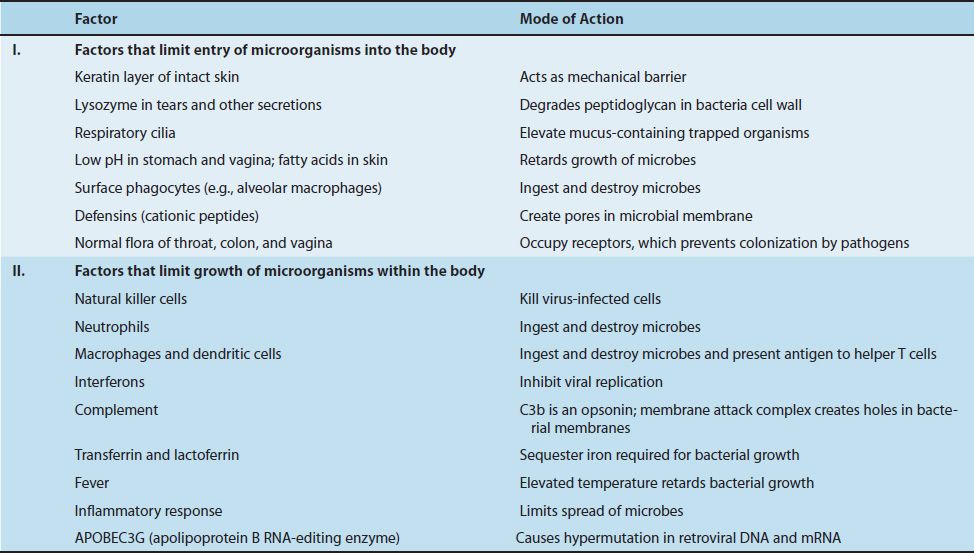
Innate immunity is resistance that exists prior to exposure to the microbe (antigen). It is nonspecific and includes host defenses such as barriers to infectious agents (e.g., skin and mucous membranes), certain cells (e.g., natural killer cells), and certain proteins (e.g., the complement cascade and interferons) and involves processes such as phagocytosis and inflammation (Table 57–4). Innate immunity does not improve after exposure to the organism, in contrast to acquired immunity, which does. In addition, innate immune processes have no memory, whereas acquired immunity is characterized by long-term memory.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


