Growth and Development
LANGUAGE OF SCIENCE
 Before reading the chapter, say each of these terms out loud. This will help you avoid stumbling over them as you read.
Before reading the chapter, say each of these terms out loud. This will help you avoid stumbling over them as you read.
[acro- top or tip, -some body]
[adolesc- grow up, -ence state]
[amnio- fetal membrane, -ic relating to, cav- hollow, -ity state]
[corona crown, radiata radiant (with rays)]
[bio- life, –log- words (study of), -y activity]
[diplo- twofold, -oid of or like]
[fertil- fruitful, -ation process]
[gran- grain, -ul- little, -osa relating to, cell storeroom]
[haplo- single, -oid of or like]
[histo- tissue, -gen- produce, -esis process]
human chorionic gonadotropin (hCG)
(koh-ree-ON-ik go-nah-doh-TROH-pin)
[im- in, -planta- set or place, -ation process]
[infan- unable to speak, -y state]
[mor- mulberry, -ula little] pl., morulae
[neo- new, -nat- birth, -al relating to]
[oo- egg, -gen- produce, -esis process]
[oo- egg, -gonium offspring] pl., oogonia
[organ- instrument (organ), -gen- produce, -esis process]
[parturi- give birth, -tion process]
[placenta flat cake] pl., placentae or placentas
[post- after, -nat- birth, -al relating to]
[pre- before, -nat- birth, -al relating to]
[prim- first, -ary state, folli– bag, -cle small]
[prim- first, -ary state, germ sprout]
[prim- first, -ary state, oo- egg, -cyte cell]
[prim- first, -ary state, charassein to engrave]
[pubert- age of maturity, -y state]
[second- second, -ary relating to, folli- bag, -cle small]
[second- second, -ary relating to, oo- egg, -cyte cell]
[senesc- grow old, -ence state]
[sperm- seed, -gen- produce, -esis process]
[sperm- seed, -gonia offspring] pl., spermatogonia
[theca sheath, cell storeroom]
[therm- heat, -taxis movement or reaction]
[tropho- nourishment, -blast sprout]
[zona belt or girdle, pellucida transparent] pl., zonae pellucidae
LANGUAGE OF MEDICINE
[ab- away from, -ruptio rupture, placentae of flat cake (placenta)]
[ante- before, -nat- birth, -al relating to]
[Virginia Apgar American physician]
[arteri- vessel (artery), -sclero- harden, -osis condition]
[ather- meal, -sclero- harden, -osis condition]
[Julius Caesar Roman emperor, -ean of]
(kon-JEN-i-tall ab-nor-MAL-i-tee)
[con- with, -genit- born, -al relating to]
[ec- out, -lamp- shine forth, -sia condition]
[em- in, -bryo- fill to bursting, –log- words (study of), -y activity]
[geronto- old age, –log- words (study of), -y activity]
[hyper- excessive, -tens- stretch or pull tight, -sion state]
[lact- milk, -ose carbohydrate (sugar), in- not, -toler- bear, -ance state]
[mast- breast, -itis inflammation]
[neo- new, -nat- born, –log- words (study of), -y activity]
[osteo- bone, -arthr- joint, -itis inflammation]
[placenta flat cake, previa gone before]
[pre- before, -lamp- shine forth, -sia condition]
pregnancy-induced hypertension (PIH)
(PREG-nan-see-in-DOOST hye-per-TEN-shun)
[hyper- excessive, -tens- stretch or pull tight, -sion state]
[presby- aging, -op- vision, -ia condition]
[pro- early, -ger- old age, -ia condition]
[puerp- childbirth, -al relating to]
[ab- away from, -or- be born, -tion process]
[terato- monster, -gen produce]
A NEW HUMAN LIFE
Production of Sex Cells
Before a new human life can begin, some preliminary processes must occur. Of utmost importance is the production of mature gametes, or sex cells, by each parent. Spermatozoa, gametes of the male parent, are produced by a process called spermatogenesis. Ova, gametes of the female parent, are produced by a process called oogenesis.
MEIOSIS
Both types of gamete production require a special form of cell division characterized by meiosis. Recall from Chapter 5 that meiosis is the orderly arrangement and distribution of chromosomes that, unlike mitosis, reduces the number of chromosomes in each daughter cell to half the number present in the parent cell.
The necessity for chromosome reduction as a preliminary step to union of the sex cells is explained by the fact that the cells of each species of living organisms contain a specific number of chromosomes. Human cells, for example, contain 23 pairs, or a total of 46 chromosomes. This total of 46 chromosomes per body cell is known as the diploid number of chromosomes. Diploid comes from the Greek diploos, meaning “twofold.” If the male and female cells united without first halving their respective chromosomes, the resulting cell would contain twice as many chromosomes as is normal for human beings. Mature ova and sperm therefore contain only 23 chromosomes, or half as many, as other human cells. This total of 23 chromosomes per sex cell is known as the haploid number of chromosomes (from the Greek haploos, meaning “single”).
Meiotic division consists of two cell divisions that take place one after the other in succession. They are referred to as meiotic division I and meiotic division II, and in both, prophase, metaphase, anaphase, and telophase occur (Figure 36-1).

In the interphase that precedes prophase I (of meiotic division I) the same events occur as take place in the interphase preceding mitotic division. Specifically, each DNA molecule replicates and thereby becomes a pair of chromatids, attached to each other only at the centromere. The term chromosome applies to any condensed chromatin with its own centromere. For simplicity’s sake, in both Figures 36-1 and 36-2, only 4 of the 46 chromosomes are shown. Notice that early in meiosis I, homologous pairs of chromosomes are moved together to form groupings called tetrads. During anaphase I the tetrads split (recall that in mitosis it is the chromosomes that split during anaphase).
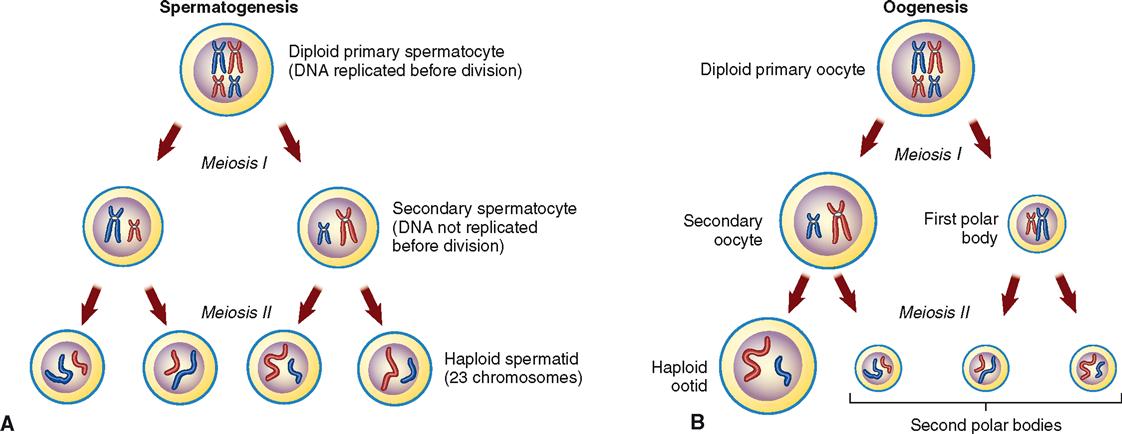
In meiosis I the phenomenon of “crossing over” occurs. During crossing over a chromatid segment of each chromosome crosses over and becomes part of the adjacent chromosome in the pair (see Figure 37-4). This is a highly significant event and is discussed in some detail in the next chapter. Because each chromatid segment consists of specific genes, the crossing over of chromatids reshuffles the genes—that is, it transfers some of them from one chromosome to another. This exchange of genetic material can add almost infinite variety to the ultimate genetic makeup of an individual.
Metaphase I follows the last stage of prophase I, and as in mitosis, the chromosomes align themselves along the equator of the spindle fibers, as Figure 36-1 shows. But in anaphase I the two chromatids that make up each chromosome do not separate from each other as they do in mitosis to form two new chromosomes out of each original one. In anaphase I, only one of each pair of chromosomes moves to each pole of the parent cell.
As you can see in Figures 36-1 and 36-2, when the parent cell divides to form two cells in meiotic division I, each daughter cell contains two chromosomes or half as many as the parent cell had. Remember that each chromosome still consists of two sister chromatids joined at the centromere. Thus the daughter cells formed by meiotic division I contain a haploid number of chromosomes, or half as many as the diploid number in the parent cell.
As you can see in Figure 36-1, meiotic division II is essentially the same as mitotic division. In both spermatogenesis and oogenesis, the second meiotic division reproduces each of the two cells formed by meiotic division I and so forms four cells, each with the haploid number of chromosomes.
SPERMATOGENESIS
Spermatogenesis is the process by which the primitive sex cells, or spermatogonia, already formed in the seminiferous tubules of a newborn baby boy can later become transformed into mature sperm, or spermatozoa. Spermatogenesis begins at about the time of puberty and usually continues throughout a man’s life. Figure 36-3 shows some of the major steps of spermatogenesis. Trace each step in this diagram with your finger as you read the following paragraph.
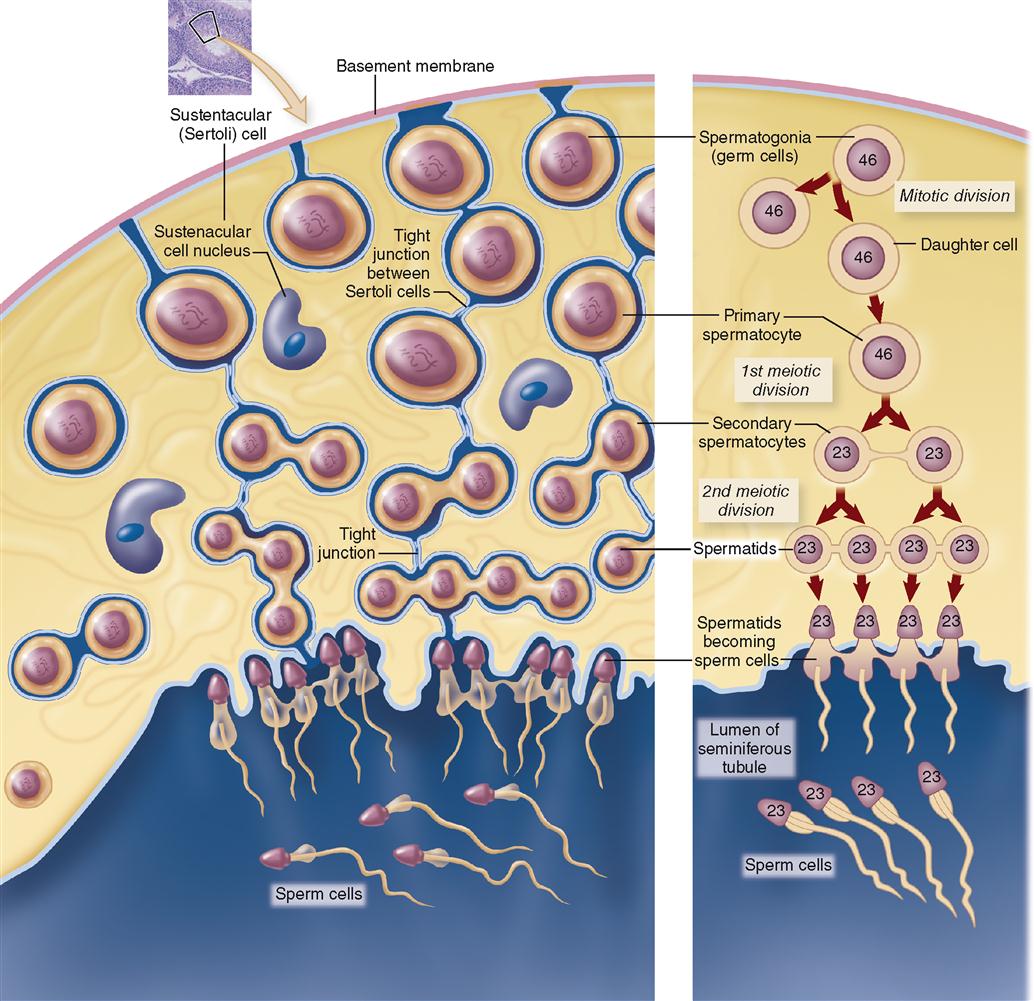
Each primary spermatocyte undergoes meiotic division I to form two secondary spermatocytes, each with a haploid number of chromosomes (23). Each secondary spermatocyte undergoes meiotic division II to form a total of four spermatids. Spermatids then differentiate to form heads and tails, eventually becoming mature spermatozoa. Thus spermatogenesis forms four spermatozoa (singular, spermatozoon)—each with only 23 chromosomes—from one primary spermatocyte that had 23 pairs, or 46 total chromosomes.
OOGENESIS
Oogenesis is the process by which primitive female sex cells, or oogonia, become mature ova. As you read through the following paragraphs, trace the steps of oogenesis in Figure 36-4.
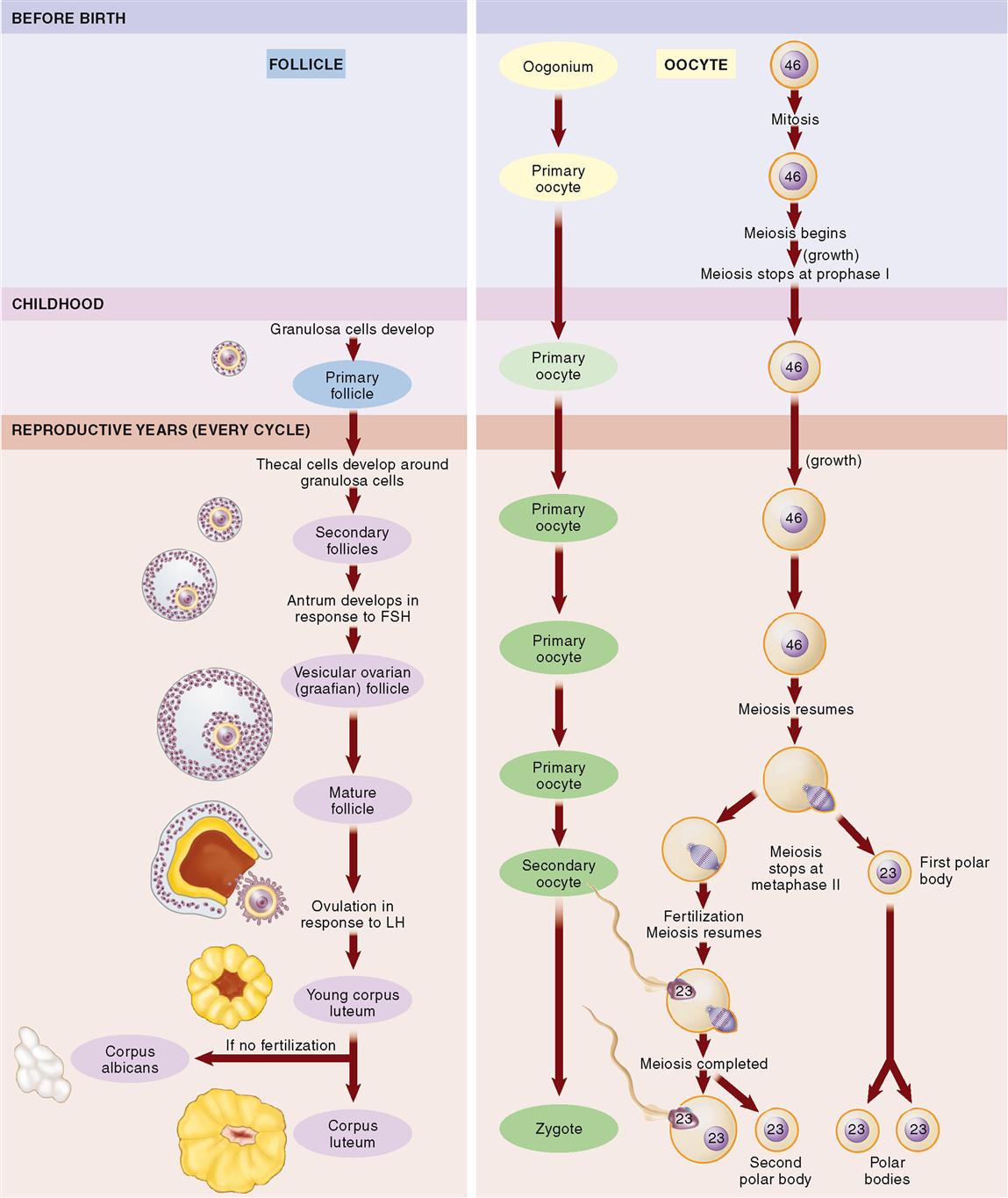
During the fetal period, oogonia in the ovaries undergo mitotic division to form primary oocytes—about a half million of them by the time a baby girl is born. Most of the primary oocytes develop to prophase I of meiosis before birth. There they stay until puberty.
What physiological advantage is gained by postponing the completion of meiotic division as an ovum matures? The surprising answer is found in Arrest of Oocyte Development online at A&P Connect.

During childhood, granulosa cells develop around each primary oocyte, forming a primary follicle. Although several thousand primary oocytes do not survive into puberty, by the time a female child reaches sexual maturity, about 400,000 primary oocytes remain.
Beginning at puberty, during each cycle, about a thousand primary oocytes resume meiosis. Their surrounding follicles begin to mature and some of the outer granulosa cells differentiate to become theca cells. Theca cells produce androgen, a steroid that is converted by granulosa cells into estrogen. At this point, the follicles are known as secondary follicles. As the secondary follicles mature, they migrate toward the surface of the ovary—sometimes in several waves during one cycle. Usually only one follicle per cycle survives and matures enough to reach the surface of the ovary, where it can be seen as a fluid-filled bump. The fluid-filled space within each mature follicle is called the antrum. A mature vesicular ovarian follicle ready to burst open from the ovary’s surface is also called a graafian follicle.
By that time, meiosis has resumed inside the primary oocyte within the mature follicle. Meiosis I produces a secondary oocyte and the first polar body (see Figure 36-2, B). Just before ovulation, meiosis again halts—this time at metaphase II. Under the influence of LH (luteinizing hormone), ovulation occurs. Ovulation, you may recall, is the release of an oocyte from a burst follicle.
Meiosis II in the released oocyte resumes only when, and if, the head of a sperm cell enters the secondary oocyte (ovum). If fertilization does not occur, then the ovum simply degenerates. If fertilization does occur, however, then meiotic division of the secondary oocyte produces a second polar body and a mature, fertilized ovum called the zygote (ZYE-goht).
Note that during oogenesis, the cytoplasm is not equally divided among the daughter cells. Of the four daughter cells produced, only one is large enough to survive. Thus each primary oocyte produces only one mature ovum, plus three tiny polar bodies. These polar bodies quickly break down and are reabsorbed into nearby cells. A total of four mature sperm cells are formed from each primary spermatocyte in spermatogenesis. This difference may be accounted for by the fact that for reproductive success, an ovum must have a huge store of cytoplasm with all of its organelles, nutrients, and regulatory molecules. In other words, nearly all the cytoplasm is conserved by the one daughter oocyte that survives.
Ovulation and Insemination
After gamete formation, the second preliminary step necessary for conception of a new individual consists of bringing the sperm and ovum into proximity with each other so that the union of the two can take place. Two processes are involved in the accomplishment of this step:
2. Insemination or expulsion of the seminal fluid from the male urethra into the female vagina. Recall from Chapter 34 that a process called capacitation occurs after ejaculation, and this enables the sperm to eventually unite with an egg. Several million sperm enter the female reproductive tract with each ejaculation of semen. By lashing movements of their flagella-like tails, assisted by various processes in the female reproductive tract, the sperm make their way into the external os of the cervix, through the cervical canal and uterine cavity, and into the uterine (fallopian) tubes.
Fertilization
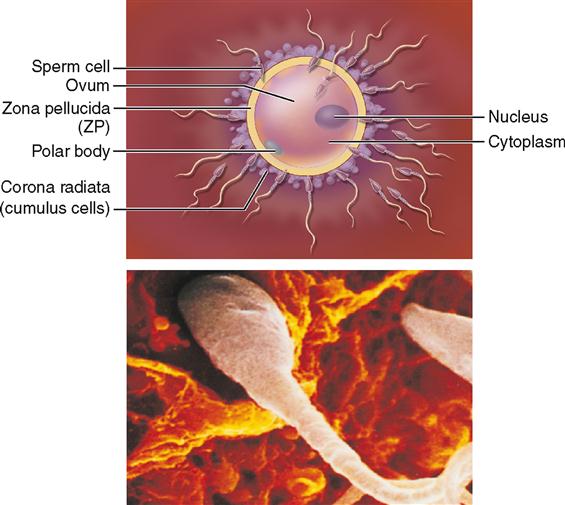
After ovulation the discharged ovum first enters the abdominopelvic cavity and then soon finds its way into the uterine (fallopian) tubes, where conception, or fertilization, may take place (Figure 36-5). Sperm cells “swim” up the uterine tubes toward the ovum. Look at the relationship of the ovary, the uterine tube, and the uterus in Figure 36-6. Recall from Chapter 35 that each uterine tube extends outward from the uterus for about 10 cm (4 inches). It then ends in the abdominal cavity near the ovary, as you can see in Figure 36-6, in an opening surrounded by fringelike processes, the fimbriae. Sperm cells that are deposited in the vagina must enter and “swim” through the uterus and then move out of the uterine cavity and through the uterine tube to meet the ovum. Fertilization most often occurs in the outer one third of the oviduct, as shown in Figure 36-6.
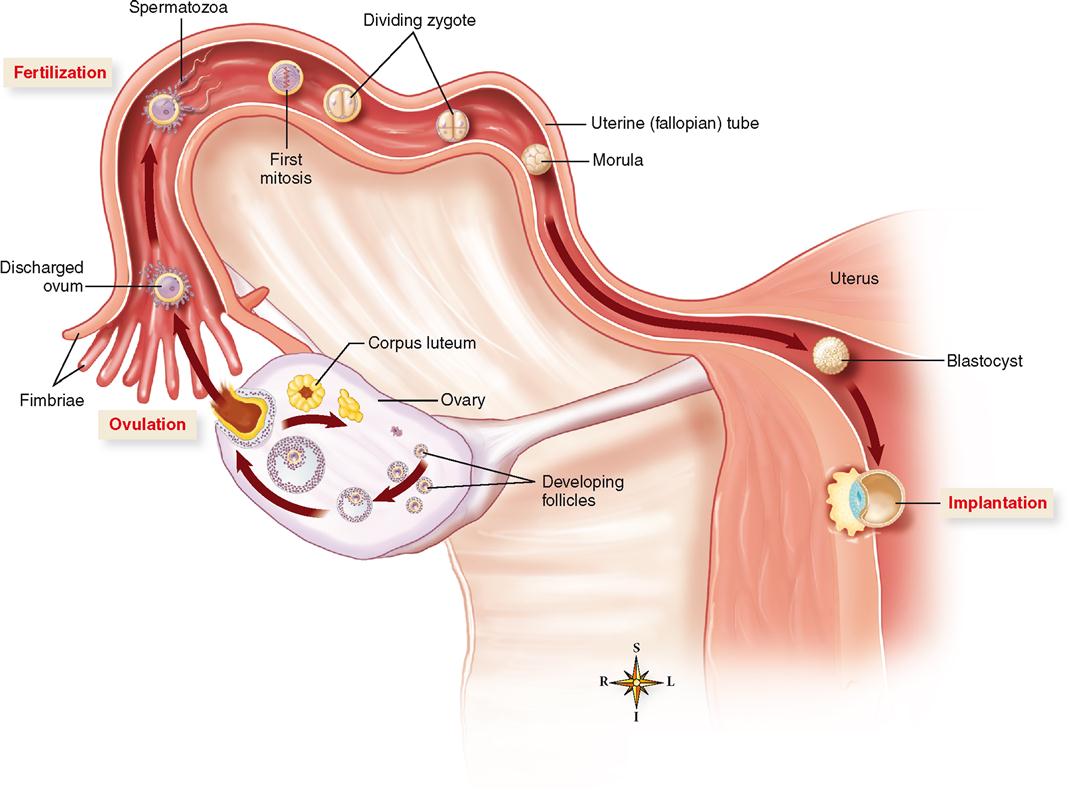
Artificial fertilization outside the body is sometimes used to enhance reproductive success. Read a brief description of this strategy and other assisted reproductive technologies (ARTs) in In Vitro Fertilization online at A&P Connect.

The process of sperm movement is assisted by mechanisms within the female reproductive tract. For example, mucous strands in the cervical canal guide the sperm on their way into the uterus. Peristaltic contractions of the female reproductive tract and ciliary movement along the lining of the uterine tubes also assist the movement of sperm. Sperm are attracted to the warmer temperatures of the uterine tubes—a process of attraction called thermotaxis. Despite all this, however, only a small fraction of the sperm deposited in the vagina ever reach the ovum. Only 50 to 100 sperm out of 250 million to 500 million sperm actually reach their target.
The ovum also takes an active role in the process of fertilization. Experiments show that the ovum and its surrounding layers actually attract nearby sperm with various regulatory molecules. Recall that such movement toward a chemical attractant is called chemotaxis. These layers around the ovum include a thick jellylike film called the zona pellucida (ZP) with an outer envelope of cumulus cells called the corona radiata. Receptor molecules on these ovum-surrounding layers bind sperm attracted to the area. Once bound to a receptor, an acrosome reaction occurs at the head of the sperm. This acrosome reaction allows the release of enzymes from the acrosome that break down the outer layers surrounding the ovum. The cumulus cells also release progesterone and other molecules that promote increased sperm motility, which aids sperm movement through the outer layers and toward the ovum.
Once the sperm reaches the surface of the ovum, the two plasma membranes fuse and the nucleus of the sperm moves inside the ovum. In addition to the sperm nucleus, RNA and protein molecules from the sperm cell also enter the egg. The RNA molecules apparently code for proteins needed early in development—thus adding to the ovum’s cellular resources. RNA molecules involved in “gene silencing” may also be released into the ovum during fertilization (see Box 5-2 on p. 120).
As soon as the head and neck of one spermatozoon fuse with the ovum (the tail degenerates), complex mechanisms in the egg are activated by sperm proteins to ensure that no more sperm enter. Specifically, sperm proteins trigger an increase in calcium concentration that causes vesicles just inside the ovum’s plasma membrane to release enzymes that inactivate the sperm receptors on the ZP. This thick film, then, becomes an impenetrable barrier sometimes called the fertilization membrane. The 23 chromosomes from the sperm nucleus combine with the 23 chromosomes already in the ovum to restore the diploid number of 46 chromosomes.
In as much as the ovum lives only a short time (probably only a day or so) after leaving the ruptured follicle, the fertilization “window” occurs around the time of ovulation. Because sperm may live up to a few days after entering the female tract, sexual intercourse any time from about 3 days before ovulation to a day or so after ovulation may result in fertilization.
The fertilized ovum, or zygote, is genetically complete; it represents the first cell of a genetically new individual. Time, nourishment, and a proper prenatal environment are all that are needed for expression of characteristics such as sex, hair, and skin color that were determined at the time of fertilization.
PRENATAL PERIOD
The prenatal period of development begins at the time of conception, or fertilization (i.e., at the moment the female ovum and the male sperm cell unite). The period of prenatal development continues until the birth of the child about 39 weeks later. The science of the development of the individual before birth is called embryology (em-bree-OL-oh-gee). It is a story of biological marvels, describing the means by which a new human life is begun and the steps by which a single microscopic cell is transformed into a complex human being.
Cleavage and Implantation
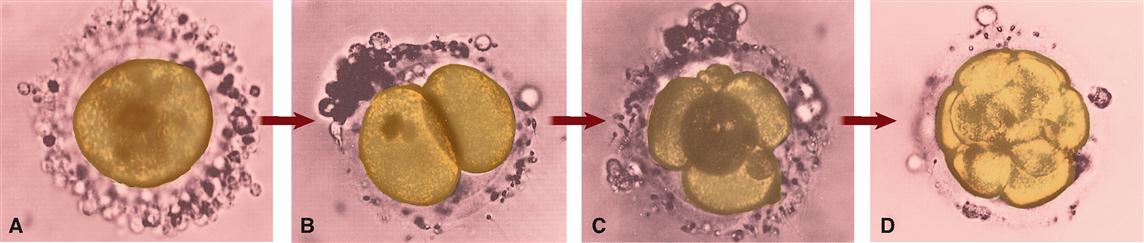
As you can see in Figure 36-6, once the zygote is formed, it immediately begins to cleave, or divide, and in about 3 days a solid mass of cells called a morula (MOR-yoo-lah) is formed. The cells of the morula begin to form an inner cavity as they continue to divide, and by the time the developing embryo reaches the uterus, it is a hollow ball of cells called a blastocyst (BLASS-toh-sist). At about 1 week after fertilization, the process of implantation begins. In about 10 days from the time of fertilization, the blastocyst is completely implanted in the uterine lining—before nutrients from the mother are available to nourish it. Of course, problems in development or implantation may occur at any stage—resulting in loss of the offspring and termination of the developmental process.
The rapid cell division taking place up to the blastocyst stage occurs with no significant increase in total mass compared with the zygote (Figure 36-7). One of the specializations of the ovum (and its surrounding layers) is its incredible store of nutrients that support this embryonic development until implantation has occurred.
Note in Figure 36-8 that the blastocyst consists of an outer layer of cells and an inner cell mass. The outer wall of the blastocyst is called the trophoblast (see Figure 36-8). As the blastocyst develops further, the inner cell mass forms a structure with two cavities called the yolk sac and amniotic (am-nee-OT-ik) cavity (Figures 36-9 and 36-10
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


