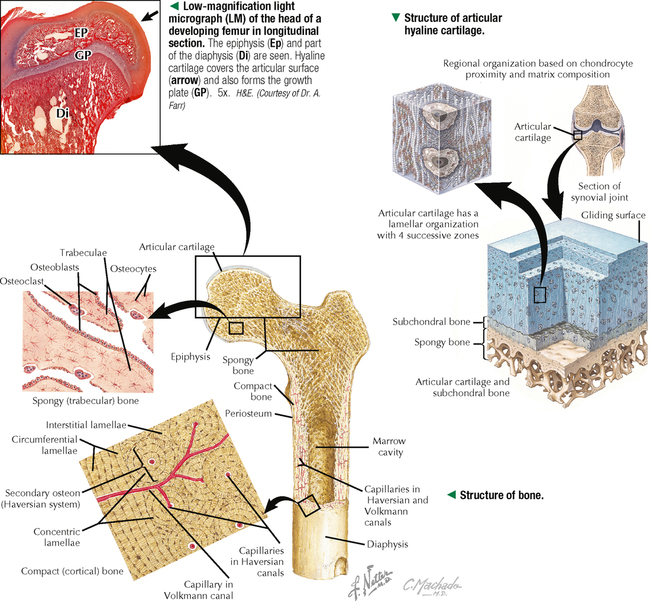
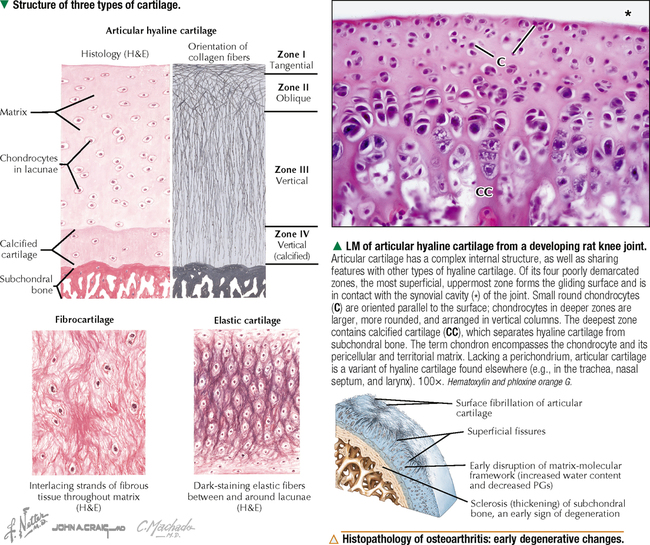
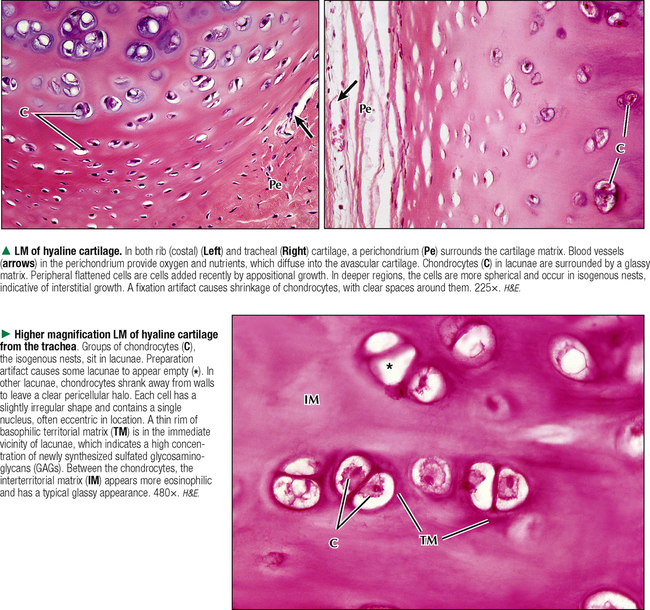
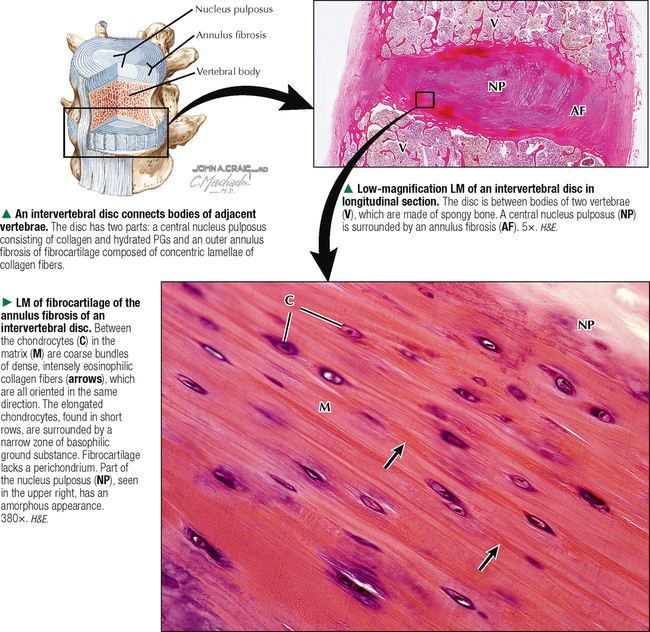
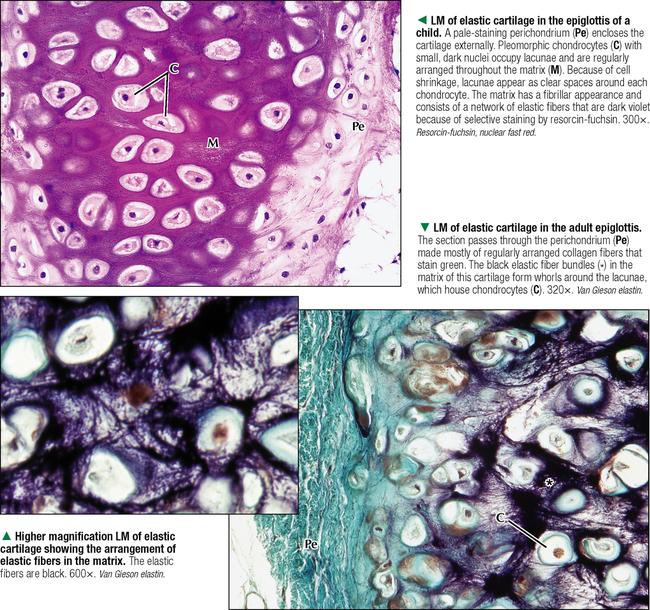
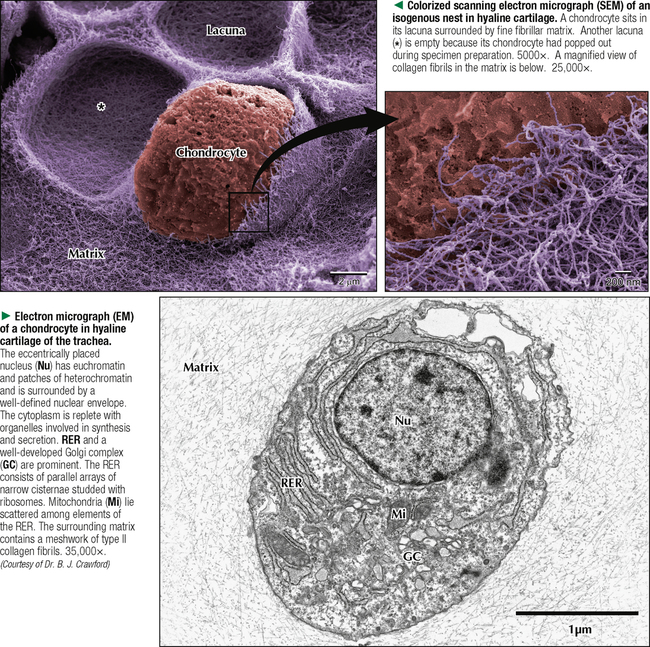
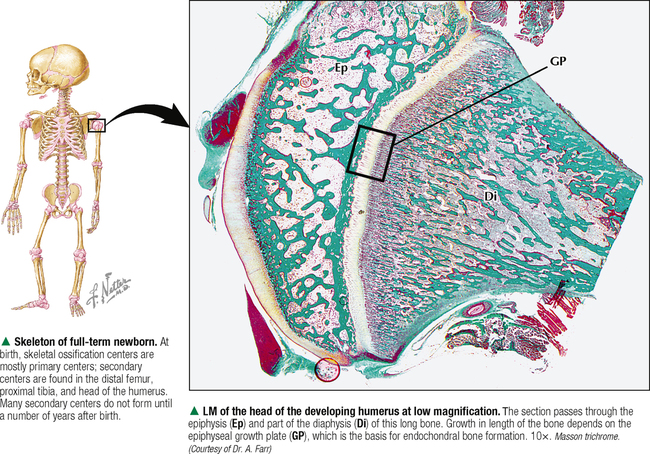
6 CARTILAGE AND BONE 6.1. Overview 6.2. Structure of Cartilage 6.3. Histology of Hyaline Cartilage 6.4. Composition of Hyaline Cartilage Matrix 6.5. Histology of Fibrocartilage 6.6. Histology of Elastic Cartilage 6.7. Ultrastructure of Chondrocytes 6.8. Overview of Bone Formation (Osteogenesis) 6.9. Intramembranous Bone Formation 6.10. Endochondral Bone Formation 6.11. Structure and Function of Growth Plates 6.12. Histology of the Growth Plate and the Metaphysis 6.13. Histology of Trabecular Bone Deposition and Resorption 6.14. Histology and Function of the Cells of Trabecular Bone 6.15. Microarchitecture of Compact Bone 6.16. Histology and Function of Spongy and Compact Bone 6.17. Structure and Function of the Periosteum 6.18. Formation and Composition of Collagen 6.19. Ultrastructure of Osteoblasts 6.20. Ultrastructure of Osteocytes 6.21. Ultrastructure and Function of Osteoclasts 6.22. Bone Fracture Repair: Early Events 6.23. Bone Fracture Repair: Intermediate and Late Events 6.24. Histology of Synovial Joints 6.25. Histology and Function of the Synovium 6.1 OVERVIEW Cartilage and bone are specialized forms of connective tissue that have critical roles in providing the skeletal framework of the body. Although they share similarities, many important differences set them apart. As with other connective tissues, they derive from embryonic mesenchyme; both consist of cells embedded in an extracellular matrix. Cartilage matrix is a firm yet resilient gel, with physical attributes of a plastic; it is not as rigid as matrix of bone and gives cartilage a solid, firm consistency. Cartilage matrix is highly hydrated, being 70%-75% water. The rest of the matrix is composed of collagen (15%-20%), for tensile strength, and proteoglycans (PGs) (2%-10%), for resilience. Cartilage provides structural support for soft tissues and a sliding area for joints and allows for growth in long bone length. Cartilage performs diverse and varied functions, but it lacks attributes of most other tissues: it is avascular and has no nerve or lymphatic supply. Bone is the calcified component of the skeleton, which in the human comprises 206 individual bones. The matrix of bone, as a rigid connective tissue, consists of collagen embedded in a ground substance on which is deposited a complex inorganic mineral, hydroxyapatite. As a tissue, compared with cartilage, bone has a higher metabolic rate, is richly vascularized, and receives up to 10% of cardiac output. Bone has good regenerative potential for self-repair throughout life, whereas cartilage has a very limited capacity for regeneration in response to traumatic injury or disease. 6.2 STRUCTURE OF CARTILAGE The three types of cartilage—hyaline cartilage, elastic cartilage, and fibrocartilage—differ mostly in histologic appearance and properties of extracellular matrix. Hyaline cartilage, the most common and characteristic type, has a matrix with a translucent, glassy appearance because the refractive index of its collagen is similar to that of the ground substance in which it is embedded. In the fetus, hyaline cartilage forms a provisional skeleton, which is replaced by bone during endochondral bone formation. Soon after birth and up to adolescence, hyaline cartilage is an integral component of epiphyseal growth plates, which control the growth and shape of long bones. In addition, hyaline cartilage lines articular surfaces of synovial joints, where it acts as a self-lubricating shock absorber with low friction properties. Hyaline cartilage also provides semirigid support to walls of some respiratory airways. Damaged hyaline cartilage is unable to be repaired because in the adult its cells—chondrocytes—cannot undergo mitosis. Elastic cartilage contains chondrocytes embedded in a matrix dominated by elastic fibers. Firm but flexible, it contributes structural integrity to the auricle of the ear, epiglottis, and eustachian (auditory) tube and allows bending. Fibrocartilage has great tensile strength because of the number of collagen fibers in its matrix. It attaches bone to tendon and, because it has load-distributing properties, it is found in menisci of synovial joints and in intervertebral discs. Dense fibrous connective tissue replaces damaged fibrocartilage. CLINICAL POINT Osteoarthritis, the most common form of arthritis, is a major cause of long-term disability in adults in North America. It is primarily a disease of articular cartilage, its hallmarks being extracellular matrix degradation and altered chondrocyte metabolism. The disorder is associated with decreased glycosaminoglycan content of the matrix accompanied by increased water content. Enhanced matrix metalloproteinase enzyme activity appears to play a major role in matrix degradation and to participate in breakdown of both PGs and collagen. Loss of cartilage leads to bone-on-bone contact in synovial joints with rapid deterioration of movement and function. 6.3 HISTOLOGY OF HYALINE CARTILAGE Except where hyaline cartilage serves as articular cartilage and is exposed to synovial fluid, it is enclosed by a layer of dense connective tissue—the perichondrium—which is essential for cartilage growth. This connective tissue investment is rich in fibroblasts, undifferentiated mesenchymal cells, blood vessels, and nerves. During growth, the perichondrium consists of an inner chondrogenic layer surrounded by a fibrous layer. In the embryo, hyaline cartilage arises from loose connective tissue when the oxygen supply is low, whereas bone arises from the same tissue when oxygen is plentiful. Chondrocytes are flattened near the perichondrium and more round in deeper regions. Chondrocytes in hyaline cartilage are often arranged in pairs or groups of four to six. Cells of a group are known as an isogenous nest because they are progeny of a single chondrocyte during development. Type II collagen fibers, water, and ground substance constitute the matrix. Collagen fibers impart eosinophilia to the matrix but are not visible by light microscopy because their refractive index is similar to that of ground substance. Basophilia of this cartilage is due to the presence of glycosaminoglycans (GAGs), such as chondroitin sulfate, in ground substance. The basophilic, metachromatic matrix immediately surrounding chondrocytes is the territorial matrix, which is rich in sulfated GAGs but contains few collagen fibers. A paler, less basophilic interterritorial matrix between chondrocytes is older synthetically and contains a fine basket-like network of type II collagen fibers. 6.4 COMPOSITION OF HYALINE CARTILAGE MATRIX Chondrocytes of hyaline cartilage are highly specialized cells that synthesize and maintain all components of the extracellular matrix. The matrix has a unique, highly ordered molecular organization. Depending on age and location of the cartilage, 60%-70% of its wet weight is water. Water and inorganic salts give cartilage its resilience and lubricating capabilities. Remaining constituents are structural macromolecules: collagens, PGs, and noncollagenous proteins. Of the dry weight of cartilage matrix, 40%-70% is collagen. Type II collagen accounts for 90%-95% of the collagen in hyaline cartilage and forms a fibrillar meshwork that mainly provides tensile strength and shape. At least two other types—IX and XI—help stabilize the network of type II fibrils. PGs in the matrix are negatively charged and hold large amounts of positively charged water ions. PGs are composed of a core protein with complex carbohydrates, known as GAGs, that radiate from the core and resemble bristles of a brush. GAGs consist of repeating, negatively charged disaccharide units of various lengths and may be sulfated or not. Sulfated PGs, mainly chondroitin sulfate, dermatan sulfate, and keratan sulfate, in turn attach noncovalently to hyaluronic acid, a nonsulfated GAG, to form large PG aggregates known as aggrecans. Interactions of aggrecans, water, and collagen fibril network give cartilage its resistance to compression (stiffness) and resilience. Chondrocytes scattered throughout the matrix attach via transmembrane proteins to the macromolecular framework they synthesize. 6.5 HISTOLOGY OF FIBROCARTILAGE Fibrocartilage is found in the symphysis pubis, the annulus fibrosis of intervertebral discs, and at points of attachment of tendons to bone. It is a mixture between dense regular connective tissue (similar in many respects to tendon or ligament) and hyaline cartilage. It combines the tensile strength, firmness, and durability of tendon with resistance to compression of cartilage. In contrast to other types of cartilage, fibrocartilage lacks a distinct perichondrium, which blends imperceptibly with surrounding connective tissue or hyaline cartilage. Its matrix is intensely eosinophilic because numerous collagen fibers are present. Arranged in parallel bundles, often in line with the direction of pull or stress applied, they give a characteristic fibrous appearance to the matrix, which is readily seen in routine histologic preparations. The matrix contains a minimal amount of amorphous ground substance, which is usually seen at boundaries of lacunae, where it is slightly basophilic or stains positively for periodic acid–Schiff (PAS). Chondrocytes are thinly dispersed in the matrix and are arranged in short, parallel rows between collagen fiber bundles. In contrast to hyaline cartilage, with type II collagen in its matrix, fibrocartilage is composed of type I collagen. Fibrocartilage initially forms from dense connective tissue that is rich in fibroblasts, some of which differentiate into chondrocytes. Thus, a mixture of chondrocytes and fibroblasts is characteristic of mature fibrocartilage. At any location, damaged hyaline or elastic cartilage is repaired via formation of fibrocartilage. 6.6 HISTOLOGY OF ELASTIC CARTILAGE Fresh elastic cartilage appears more opaque and yellow than hyaline cartilage because of abundant elastic fibers in its matrix. Elastic cartilage is resilient, easily returning to its original shape after bending or distortion, and has more flexibility and elasticity than other cartilage types. Its matrix contains a dense, interwoven network of elastic fibers embedded in a small amount of amorphous extracellular ground substance. This network is denser in the interior than at the periphery. The spherical chondrocytes, which sit in lacunae, appear similar to chondrocytes of hyaline cartilage, except that they are more closely packed and often found singly in the lacunae (only a few isogenous nests are present). The high refractive index of elastic fibers imparts a lightly eosinophilic staining pattern in conventional preparations. With methods that stain selectively for elastin, the branching and anastomosing of the elastic fibers are seen more clearly. The matrix also contains a small number of type II collagen fibers that are masked by ground substance and intermingle with the more abundant elastic fibers. Like hyaline cartilage (other than that on articular surfaces of joints), elastic cartilage is enveloped by a perichondrium. Blood vessels and lymphatics in the perichondrium do not penetrate the cartilage interior. Elastic cartilage undergoes either appositional growth, from the perichondrium, or interstitial growth, by chondrocyte mitosis. In contrast to other types of cartilage, elastic cartilage does not calcify with age. 6.7 ULTRASTRUCTURE OF CHONDROCYTES Ultrastructural features of chondrocytes reflect function: synthesis and secretion of all components of the extracellular matrix. These features are similar to those of other cells such as fibroblasts and osteoblasts that synthesize and secrete proteins such as collagen as well as carbohydrates that make up ground substance. In optimal electron microscopic preparations, a chondrocyte completely fills its lacuna. Its plasma membrane has small, irregular extensions, or footlets, that project at various points around the periphery. In active cells, a single, irregularly ovoid nucleus is mostly euchromatic with peripheral clumps of heterochromatin close to the nuclear envelope. The cytoplasm contains abundant free ribosomes and well-developed rough endoplasmic reticulum (RER) with dilated cisternae. The prominent juxtanuclear Golgi complex contains expanded saccules and vacuoles of various sizes. Mitochondria are sporadic, which most likely reflects their energy utilization from glycolytic, or anaerobic, rather than aerobic mechanisms. Fat droplets and extensive clusters of glycogen are present, especially in mature, less active cells. Delicate type II collagen fibrils dispersed in the matrix have a faintly cross-banded appearance at high magnification but lack the 64-nm periodicity characteristic of type I collagen in other sites. Type II fibrils form a loose three-dimensional network throughout the matrix and do not assemble into coarse bundles. 6.8 OVERVIEW OF BONE FORMATION (OSTEOGENESIS) Bone formation, or osteogenesis, is an astounding, complex series of interrelated, simultaneously occurring processes: cell migration, mitosis, differentiation, modulation, synthesis, secretion, extracellular mineralization, and resorption. Development of the skeleton begins in the early embryo and fetus, with growth continuing after birth and up to adolescence. Bones are formed by either intramembranous or endochondral ossification. These two types of osteogenesis refer only to the initial environment in which a bone forms—whether bone forms on the basis of a cartilage model (endochondral) or not (intramembranous)—not the microscopic structure of completely developed bone. Intramembranous ossification occurs in areas of ordinary mesenchyme where osteoblasts, or bone-forming cells, differentiate directly within richly vascularized mesenchymal connective tissue. Flat bones of the cranium, part of the mandible, and the clavicles develop in this way. Most long bones (of the extremities), vertebral column, ribs, and pelvis develop by endochondral ossification in preexisting hyaline cartilage models. Here, mesenchymal cells differentiate into chondrocytes. A cartilage template is modified to facilitate mineralization, vascular invasion, and replacement by bone. Both kinds of osteogenesis have the same mechanism of bone matrix deposition and mineralization by first producing spongy (cancellous, or trabecular) bone. Much of this bone later develops into dense (compact, or cortical) bone. After bone forms, it remains dynamic throughout life to allow growth and serve as a source of mineral ions for homeostasis. CLINICAL POINT Paget disease (osteodystrophia deformans), a chronic disorder of excessive bone resorption followed by abnormal bone formation, results in enlarged, misshapen, brittle bones that are prone to fracture. Remodeling and increased bone size cause stretching of the periosteum and nerve endings, leading to pain. Most frequently seen in middle-aged to elderly men, the disease was first described in 1877 by British surgeon Sir James Paget. The most commonly affected bones are the humerus, clavicle, femur, vertebrae, and pelvis. On biopsy, the appearance of small, irregular fragments of bone matrix united in a jigsaw or mosaic pattern is the histologic hallmark. Its etiology is unknown, but slow viral infection and genetic mutations Only gold members can continue reading. Log In or Register to continue Share this: Share on X (Opens in new window) X Share on Facebook (Opens in new window) Facebook Like this:Like Loading… Related Related posts: CARDIOVASCULAR SYSTEM RESPIRATORY SYSTEM SPECIAL SENSES FEMALE REPRODUCTIVE SYSTEM Stay updated, free articles. Join our Telegram channel Join Tags: Netters Essential Histology Jun 18, 2016 | Posted by admin in HISTOLOGY | Comments Off on CARTILAGE AND BONE Full access? Get Clinical Tree
6 CARTILAGE AND BONE 6.1. Overview 6.2. Structure of Cartilage 6.3. Histology of Hyaline Cartilage 6.4. Composition of Hyaline Cartilage Matrix 6.5. Histology of Fibrocartilage 6.6. Histology of Elastic Cartilage 6.7. Ultrastructure of Chondrocytes 6.8. Overview of Bone Formation (Osteogenesis) 6.9. Intramembranous Bone Formation 6.10. Endochondral Bone Formation 6.11. Structure and Function of Growth Plates 6.12. Histology of the Growth Plate and the Metaphysis 6.13. Histology of Trabecular Bone Deposition and Resorption 6.14. Histology and Function of the Cells of Trabecular Bone 6.15. Microarchitecture of Compact Bone 6.16. Histology and Function of Spongy and Compact Bone 6.17. Structure and Function of the Periosteum 6.18. Formation and Composition of Collagen 6.19. Ultrastructure of Osteoblasts 6.20. Ultrastructure of Osteocytes 6.21. Ultrastructure and Function of Osteoclasts 6.22. Bone Fracture Repair: Early Events 6.23. Bone Fracture Repair: Intermediate and Late Events 6.24. Histology of Synovial Joints 6.25. Histology and Function of the Synovium 6.1 OVERVIEW Cartilage and bone are specialized forms of connective tissue that have critical roles in providing the skeletal framework of the body. Although they share similarities, many important differences set them apart. As with other connective tissues, they derive from embryonic mesenchyme; both consist of cells embedded in an extracellular matrix. Cartilage matrix is a firm yet resilient gel, with physical attributes of a plastic; it is not as rigid as matrix of bone and gives cartilage a solid, firm consistency. Cartilage matrix is highly hydrated, being 70%-75% water. The rest of the matrix is composed of collagen (15%-20%), for tensile strength, and proteoglycans (PGs) (2%-10%), for resilience. Cartilage provides structural support for soft tissues and a sliding area for joints and allows for growth in long bone length. Cartilage performs diverse and varied functions, but it lacks attributes of most other tissues: it is avascular and has no nerve or lymphatic supply. Bone is the calcified component of the skeleton, which in the human comprises 206 individual bones. The matrix of bone, as a rigid connective tissue, consists of collagen embedded in a ground substance on which is deposited a complex inorganic mineral, hydroxyapatite. As a tissue, compared with cartilage, bone has a higher metabolic rate, is richly vascularized, and receives up to 10% of cardiac output. Bone has good regenerative potential for self-repair throughout life, whereas cartilage has a very limited capacity for regeneration in response to traumatic injury or disease. 6.2 STRUCTURE OF CARTILAGE The three types of cartilage—hyaline cartilage, elastic cartilage, and fibrocartilage—differ mostly in histologic appearance and properties of extracellular matrix. Hyaline cartilage, the most common and characteristic type, has a matrix with a translucent, glassy appearance because the refractive index of its collagen is similar to that of the ground substance in which it is embedded. In the fetus, hyaline cartilage forms a provisional skeleton, which is replaced by bone during endochondral bone formation. Soon after birth and up to adolescence, hyaline cartilage is an integral component of epiphyseal growth plates, which control the growth and shape of long bones. In addition, hyaline cartilage lines articular surfaces of synovial joints, where it acts as a self-lubricating shock absorber with low friction properties. Hyaline cartilage also provides semirigid support to walls of some respiratory airways. Damaged hyaline cartilage is unable to be repaired because in the adult its cells—chondrocytes—cannot undergo mitosis. Elastic cartilage contains chondrocytes embedded in a matrix dominated by elastic fibers. Firm but flexible, it contributes structural integrity to the auricle of the ear, epiglottis, and eustachian (auditory) tube and allows bending. Fibrocartilage has great tensile strength because of the number of collagen fibers in its matrix. It attaches bone to tendon and, because it has load-distributing properties, it is found in menisci of synovial joints and in intervertebral discs. Dense fibrous connective tissue replaces damaged fibrocartilage. CLINICAL POINT Osteoarthritis, the most common form of arthritis, is a major cause of long-term disability in adults in North America. It is primarily a disease of articular cartilage, its hallmarks being extracellular matrix degradation and altered chondrocyte metabolism. The disorder is associated with decreased glycosaminoglycan content of the matrix accompanied by increased water content. Enhanced matrix metalloproteinase enzyme activity appears to play a major role in matrix degradation and to participate in breakdown of both PGs and collagen. Loss of cartilage leads to bone-on-bone contact in synovial joints with rapid deterioration of movement and function. 6.3 HISTOLOGY OF HYALINE CARTILAGE Except where hyaline cartilage serves as articular cartilage and is exposed to synovial fluid, it is enclosed by a layer of dense connective tissue—the perichondrium—which is essential for cartilage growth. This connective tissue investment is rich in fibroblasts, undifferentiated mesenchymal cells, blood vessels, and nerves. During growth, the perichondrium consists of an inner chondrogenic layer surrounded by a fibrous layer. In the embryo, hyaline cartilage arises from loose connective tissue when the oxygen supply is low, whereas bone arises from the same tissue when oxygen is plentiful. Chondrocytes are flattened near the perichondrium and more round in deeper regions. Chondrocytes in hyaline cartilage are often arranged in pairs or groups of four to six. Cells of a group are known as an isogenous nest because they are progeny of a single chondrocyte during development. Type II collagen fibers, water, and ground substance constitute the matrix. Collagen fibers impart eosinophilia to the matrix but are not visible by light microscopy because their refractive index is similar to that of ground substance. Basophilia of this cartilage is due to the presence of glycosaminoglycans (GAGs), such as chondroitin sulfate, in ground substance. The basophilic, metachromatic matrix immediately surrounding chondrocytes is the territorial matrix, which is rich in sulfated GAGs but contains few collagen fibers. A paler, less basophilic interterritorial matrix between chondrocytes is older synthetically and contains a fine basket-like network of type II collagen fibers. 6.4 COMPOSITION OF HYALINE CARTILAGE MATRIX Chondrocytes of hyaline cartilage are highly specialized cells that synthesize and maintain all components of the extracellular matrix. The matrix has a unique, highly ordered molecular organization. Depending on age and location of the cartilage, 60%-70% of its wet weight is water. Water and inorganic salts give cartilage its resilience and lubricating capabilities. Remaining constituents are structural macromolecules: collagens, PGs, and noncollagenous proteins. Of the dry weight of cartilage matrix, 40%-70% is collagen. Type II collagen accounts for 90%-95% of the collagen in hyaline cartilage and forms a fibrillar meshwork that mainly provides tensile strength and shape. At least two other types—IX and XI—help stabilize the network of type II fibrils. PGs in the matrix are negatively charged and hold large amounts of positively charged water ions. PGs are composed of a core protein with complex carbohydrates, known as GAGs, that radiate from the core and resemble bristles of a brush. GAGs consist of repeating, negatively charged disaccharide units of various lengths and may be sulfated or not. Sulfated PGs, mainly chondroitin sulfate, dermatan sulfate, and keratan sulfate, in turn attach noncovalently to hyaluronic acid, a nonsulfated GAG, to form large PG aggregates known as aggrecans. Interactions of aggrecans, water, and collagen fibril network give cartilage its resistance to compression (stiffness) and resilience. Chondrocytes scattered throughout the matrix attach via transmembrane proteins to the macromolecular framework they synthesize. 6.5 HISTOLOGY OF FIBROCARTILAGE Fibrocartilage is found in the symphysis pubis, the annulus fibrosis of intervertebral discs, and at points of attachment of tendons to bone. It is a mixture between dense regular connective tissue (similar in many respects to tendon or ligament) and hyaline cartilage. It combines the tensile strength, firmness, and durability of tendon with resistance to compression of cartilage. In contrast to other types of cartilage, fibrocartilage lacks a distinct perichondrium, which blends imperceptibly with surrounding connective tissue or hyaline cartilage. Its matrix is intensely eosinophilic because numerous collagen fibers are present. Arranged in parallel bundles, often in line with the direction of pull or stress applied, they give a characteristic fibrous appearance to the matrix, which is readily seen in routine histologic preparations. The matrix contains a minimal amount of amorphous ground substance, which is usually seen at boundaries of lacunae, where it is slightly basophilic or stains positively for periodic acid–Schiff (PAS). Chondrocytes are thinly dispersed in the matrix and are arranged in short, parallel rows between collagen fiber bundles. In contrast to hyaline cartilage, with type II collagen in its matrix, fibrocartilage is composed of type I collagen. Fibrocartilage initially forms from dense connective tissue that is rich in fibroblasts, some of which differentiate into chondrocytes. Thus, a mixture of chondrocytes and fibroblasts is characteristic of mature fibrocartilage. At any location, damaged hyaline or elastic cartilage is repaired via formation of fibrocartilage. 6.6 HISTOLOGY OF ELASTIC CARTILAGE Fresh elastic cartilage appears more opaque and yellow than hyaline cartilage because of abundant elastic fibers in its matrix. Elastic cartilage is resilient, easily returning to its original shape after bending or distortion, and has more flexibility and elasticity than other cartilage types. Its matrix contains a dense, interwoven network of elastic fibers embedded in a small amount of amorphous extracellular ground substance. This network is denser in the interior than at the periphery. The spherical chondrocytes, which sit in lacunae, appear similar to chondrocytes of hyaline cartilage, except that they are more closely packed and often found singly in the lacunae (only a few isogenous nests are present). The high refractive index of elastic fibers imparts a lightly eosinophilic staining pattern in conventional preparations. With methods that stain selectively for elastin, the branching and anastomosing of the elastic fibers are seen more clearly. The matrix also contains a small number of type II collagen fibers that are masked by ground substance and intermingle with the more abundant elastic fibers. Like hyaline cartilage (other than that on articular surfaces of joints), elastic cartilage is enveloped by a perichondrium. Blood vessels and lymphatics in the perichondrium do not penetrate the cartilage interior. Elastic cartilage undergoes either appositional growth, from the perichondrium, or interstitial growth, by chondrocyte mitosis. In contrast to other types of cartilage, elastic cartilage does not calcify with age. 6.7 ULTRASTRUCTURE OF CHONDROCYTES Ultrastructural features of chondrocytes reflect function: synthesis and secretion of all components of the extracellular matrix. These features are similar to those of other cells such as fibroblasts and osteoblasts that synthesize and secrete proteins such as collagen as well as carbohydrates that make up ground substance. In optimal electron microscopic preparations, a chondrocyte completely fills its lacuna. Its plasma membrane has small, irregular extensions, or footlets, that project at various points around the periphery. In active cells, a single, irregularly ovoid nucleus is mostly euchromatic with peripheral clumps of heterochromatin close to the nuclear envelope. The cytoplasm contains abundant free ribosomes and well-developed rough endoplasmic reticulum (RER) with dilated cisternae. The prominent juxtanuclear Golgi complex contains expanded saccules and vacuoles of various sizes. Mitochondria are sporadic, which most likely reflects their energy utilization from glycolytic, or anaerobic, rather than aerobic mechanisms. Fat droplets and extensive clusters of glycogen are present, especially in mature, less active cells. Delicate type II collagen fibrils dispersed in the matrix have a faintly cross-banded appearance at high magnification but lack the 64-nm periodicity characteristic of type I collagen in other sites. Type II fibrils form a loose three-dimensional network throughout the matrix and do not assemble into coarse bundles. 6.8 OVERVIEW OF BONE FORMATION (OSTEOGENESIS) Bone formation, or osteogenesis, is an astounding, complex series of interrelated, simultaneously occurring processes: cell migration, mitosis, differentiation, modulation, synthesis, secretion, extracellular mineralization, and resorption. Development of the skeleton begins in the early embryo and fetus, with growth continuing after birth and up to adolescence. Bones are formed by either intramembranous or endochondral ossification. These two types of osteogenesis refer only to the initial environment in which a bone forms—whether bone forms on the basis of a cartilage model (endochondral) or not (intramembranous)—not the microscopic structure of completely developed bone. Intramembranous ossification occurs in areas of ordinary mesenchyme where osteoblasts, or bone-forming cells, differentiate directly within richly vascularized mesenchymal connective tissue. Flat bones of the cranium, part of the mandible, and the clavicles develop in this way. Most long bones (of the extremities), vertebral column, ribs, and pelvis develop by endochondral ossification in preexisting hyaline cartilage models. Here, mesenchymal cells differentiate into chondrocytes. A cartilage template is modified to facilitate mineralization, vascular invasion, and replacement by bone. Both kinds of osteogenesis have the same mechanism of bone matrix deposition and mineralization by first producing spongy (cancellous, or trabecular) bone. Much of this bone later develops into dense (compact, or cortical) bone. After bone forms, it remains dynamic throughout life to allow growth and serve as a source of mineral ions for homeostasis. CLINICAL POINT Paget disease (osteodystrophia deformans), a chronic disorder of excessive bone resorption followed by abnormal bone formation, results in enlarged, misshapen, brittle bones that are prone to fracture. Remodeling and increased bone size cause stretching of the periosteum and nerve endings, leading to pain. Most frequently seen in middle-aged to elderly men, the disease was first described in 1877 by British surgeon Sir James Paget. The most commonly affected bones are the humerus, clavicle, femur, vertebrae, and pelvis. On biopsy, the appearance of small, irregular fragments of bone matrix united in a jigsaw or mosaic pattern is the histologic hallmark. Its etiology is unknown, but slow viral infection and genetic mutations Only gold members can continue reading. Log In or Register to continue Share this: Share on X (Opens in new window) X Share on Facebook (Opens in new window) Facebook Like this:Like Loading… Related Related posts: CARDIOVASCULAR SYSTEM RESPIRATORY SYSTEM SPECIAL SENSES FEMALE REPRODUCTIVE SYSTEM Stay updated, free articles. Join our Telegram channel Join Tags: Netters Essential Histology Jun 18, 2016 | Posted by admin in HISTOLOGY | Comments Off on CARTILAGE AND BONE Full access? Get Clinical Tree