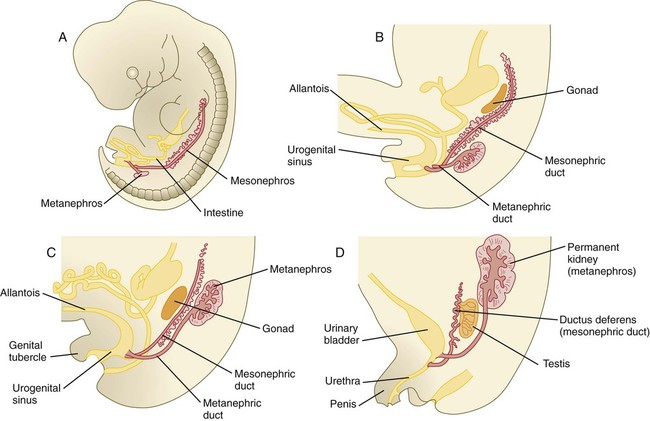
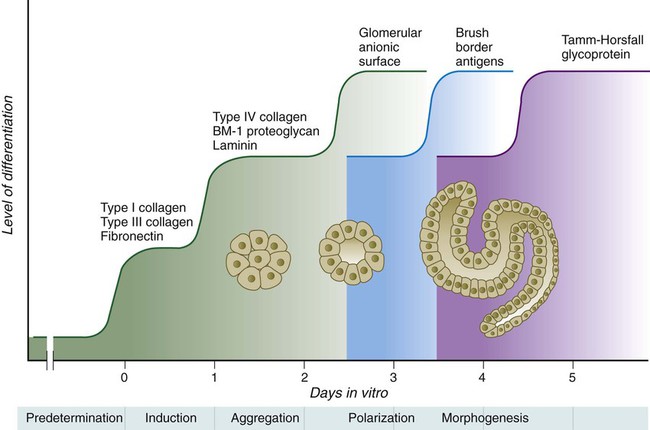
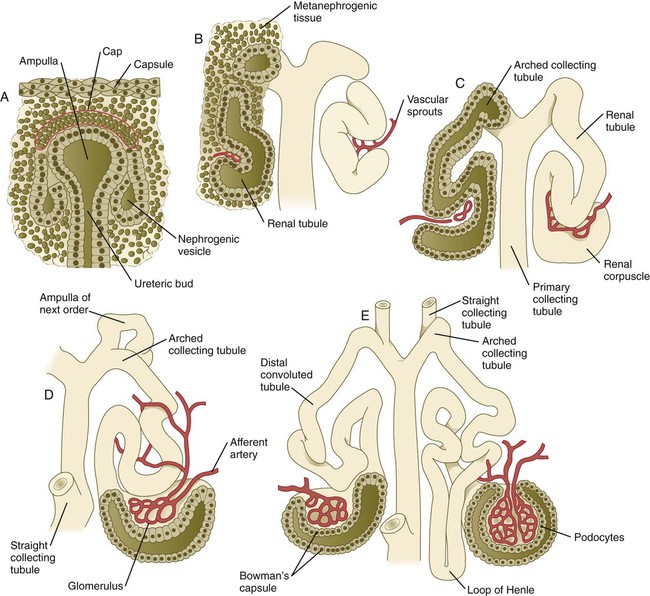
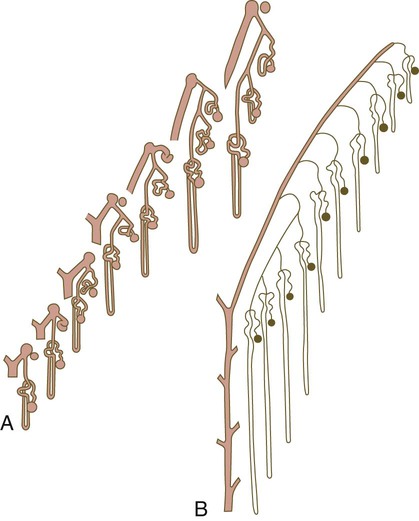
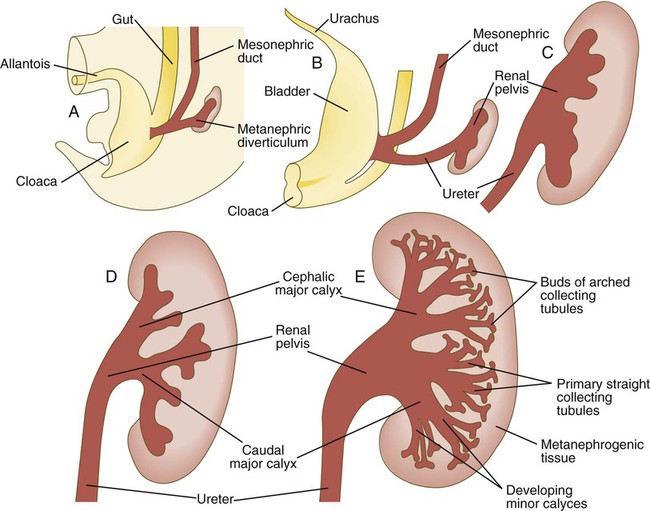
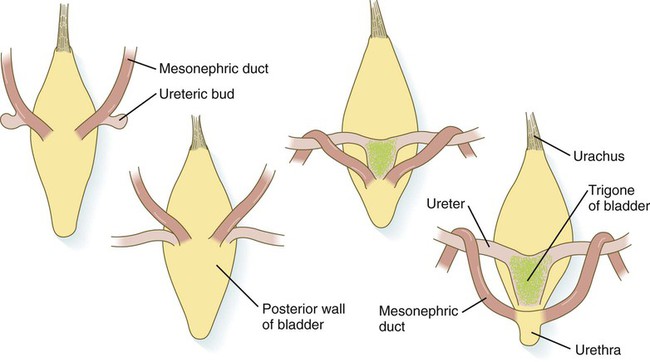
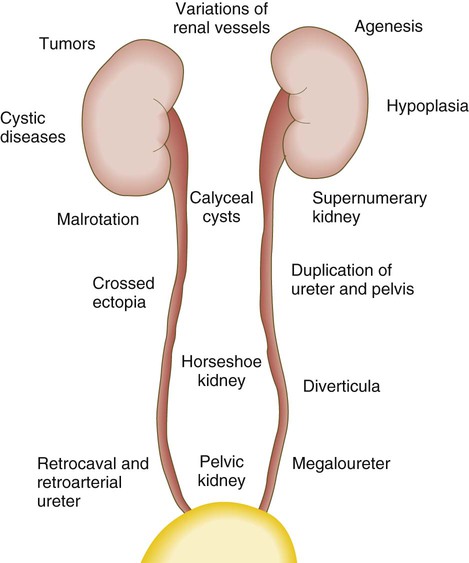
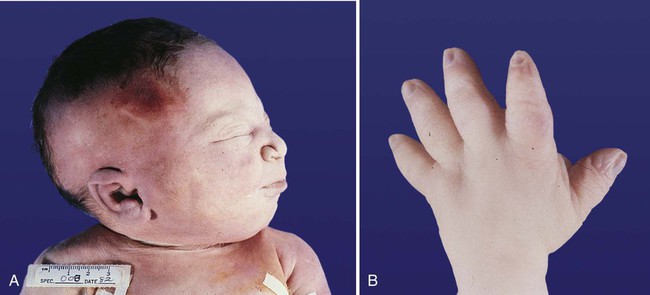
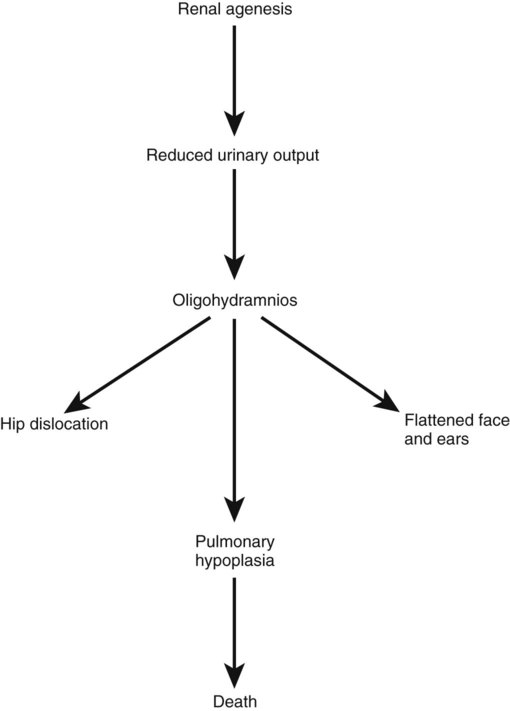
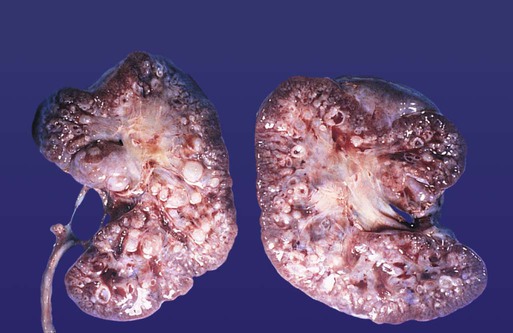
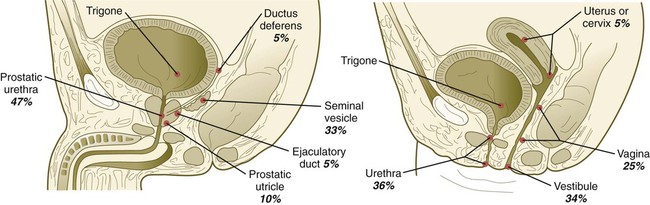
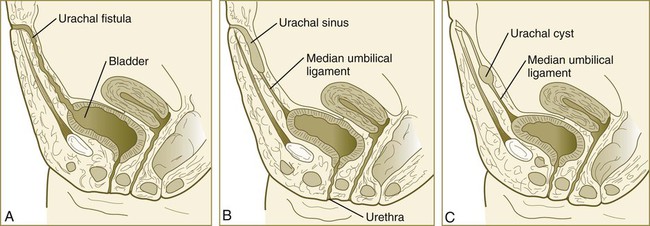
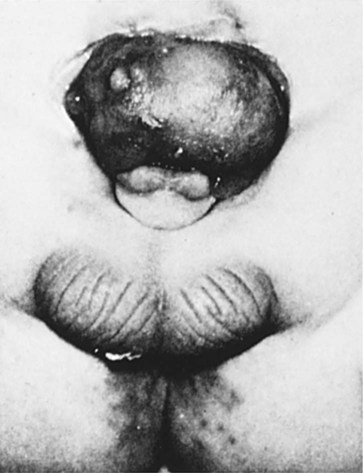
Chapter 16 The urogenital system arises from the intermediate mesoderm of the early embryo (see Fig. 6.7). Several major themes underlie the development of urinary and genital structures from this common precursor. The first is the interconnectedness of urinary and genital development, in which early components of one system are taken over by another during its later development. A second is the recapitulation during human ontogeny of kidney types (the equivalent of organ isoforms) that are terminal forms of the kidney in lower vertebrates. A third theme comprises the dependence of differentiation and the maintenance of many structures in the urogenital system on epithelial-mesenchymal interactions. Finally, the sexual differentiation of many structures passes from an indifferent stage, in which male and female differences are not readily apparent, to a male or female pathway, depending on the presence of specific promoting or inhibiting factors acting on the structure. Although phenotypic sex is genetically determined, genetic sex can be overridden by environmental factors, thus leading to a discordance between the two. Clinical Correlations 16.1 and 16.2, later in this chapter, discuss abnormalities of the urinary and genital systems, respectively. The common representation of mammalian kidney development includes three successive phases beginning with the appearance of the pronephros, the developmental homologue of the type of kidney found in only the lowest vertebrates. In human embryos, the first evidence of a urinary system consists of the appearance of a few segmentally arranged sets of epithelial cords that differentiate from the anterior intermediate mesoderm at about 22 days’ gestation. These structures are more appropriately called nephrotomes. The nephrotomes connect laterally with a pair of primary nephric (pronephric) ducts, which grow toward the cloaca (Fig. 16.1). The earliest stages in the development of the urinary system depend on the action of retinoic acid, which sets the expression limits of Hox 4-11 genes that determine the craniocaudal limits of the early urinary system. The molecular response by the intermediate mesoderm is the expression of the transcription factors Pax-2 and Pax-8, which then induce Lim-1 (Lhx-1) in the intermediate mesoderm. Lim-1 is required for the aggregation of the mesenchymal cells of the intermediate mesoderm into the primary nephric ducts. As the primary nephric ducts extend caudally, they stimulate the intermediate mesoderm to form additional segmental sets of tubules. The conversion of the mesenchymal cells of the intermediate mesoderm into epithelial tubules depends on the expression of Pax-2, and in the absence of this molecule, further development of kidney tubules does not occur. These tubules are structurally equivalent to the mesonephric tubules of fishes and amphibians. A typical mesonephric unit consists of a vascular glomerulus, which is partially surrounded by an epithelial glomerular capsule. The glomerular capsule is continuous with a contorted mesonephric tubule, which is surrounded by a mesh of capillaries (see Fig. 16.1B). Each mesonephric tubule empties separately into the continuation of the primary nephric duct, which becomes known as the mesonephric (wolffian) duct. The formation of pairs of mesonephric tubules occurs along a craniocaudal gradient. The first 4 to 6 pairs of mesonephric tubules (and the pronephric tubules) arise as outgrowths from the primary nephric ducts. Farther caudally, mesonephric tubules, up to a total of 36 to 40, take shape separately in the intermediate mesoderm slightly behind the caudal extension of the mesonephric ducts. By the end of the fourth week of gestation, the mesonephric ducts attach to the cloaca, and a continuous lumen is present throughout each. There is a difference in the developmental controls between the most cranial 4 to 6 pairs of mesonephric tubules and the remaining caudal tubules. Knockouts for the WT-1 (Wilms’ tumor suppressor) gene result in the absence of posterior mesonephric tubules, whereas the cranial tubules that bud off the pronephric duct form normally. As is the case in the formation of the metanephros (see later), WT-1 regulates the transformation from mesenchyme to epithelium during the early formation of renal (mesonephric) tubules. Very near its attachment site to the cloaca, the mesonephric duct develops an epithelial outgrowth called the ureteric bud (see Fig. 16.1A). The mesonephros is most prominent while the definitive metanephros is beginning to take shape. Although it rapidly regresses as a urinary unit after the metanephric kidneys become functional, the mesonephric ducts and some of the mesonephric tubules persist in the male and become incorporated as integral components of the genital duct system (Fig. 16.2). Development of the metanephros begins early in week 5 of gestation, when the ureteric bud (metanephric diverticulum) grows into the posterior portion of the intermediate mesoderm. Mesenchymal cells of the intermediate mesoderm condense around the metanephric diverticulum to form the metanephrogenic blastema (see Fig. 16.1C). Outgrowth of the ureteric bud from the mesonephric duct is a response to the secretion of glial cell line–derived neurotrophic factor (GDNF) by the undifferentiated mesenchyme of the metanephrogenic blastema (Fig. 16.3A). This inductive signal is bound by c-Ret, a member of the tyrosine kinase receptor superfamily, and the coreceptor Gfra-1, which are located in the plasma membranes of the epithelial cells of the early ureteric bud. The formation of GDNF in the metanephric mesenchyme is regulated by WT-1. The posterior location of the ureteric bud results from a combination of repression of GDNF expression in the more anterior regions by the actions of Slit-2/Robo-2 in the mesenchyme and Sprouty, which reduces the sensitivity of the anterior mesonephric duct to the action of GDNF. Bone morphogenetic protein (BMP) signaling in the surrounding mesoderm is also inhibitory to outgrowth of the ureteric bud, but within the metanephrogenic blastema its action is counteracted by the BMP-inhibitory actions of gremlin, which is produced within the blastema itself. The formation of individual functional tubules (nephrons) in the developing metanephros involves three mesodermal cell lineages: epithelial cells derived from the ureteric bud, mesenchymal cells of the metanephrogenic blastema, and ingrowing vascular endothelial cells. The earliest stage is the condensation of mesenchymal blastemal cells around the terminal bud of the ureteric bud (later to become the metanephric duct). The preinduced mesenchyme contains several interstitial proteins, such as types I and III collagen and fibronectin. As the mesenchymal cells condense after local induction by the branching tips of the ureteric bud, these proteins are lost and are replaced with epithelial-type proteins (type IV collagen, syndecan-1, laminin, and heparin sulfate proteoglycan), which are ultimately localized to the basement membranes (Fig. 16.4). As the terminal bud of the metanephric duct branches, each tip is surrounded by a cap of condensed mesenchyme. Soon, this cap becomes subdivided into a persisting mesenchymal cap and, at its end, a region where the mesenchyme is transforming into an epithelial nephrogenic vesicle (Fig. 16.5A). A single condensation of mesenchymal cells undergoes a defined series of stages to form a renal tubule. After a growth phase, mitotic activity within the rounded blastemal mesenchyme decreases, and the primordium of the tubule assumes a comma shape. Within the comma, a group of cells farthest from the end of the metanephric duct becomes polarized and forms a central lumen and a basal lamina on the outer surface. This marks the transformation of the induced mesenchymal cells into an epithelium—the specialized podocytes, which ultimately surround the vascular endothelium of the glomerulus. A consequence of this epithelial transformation is the formation of a slit just beneath the transforming podocyte precursors in the tubular primordium (Fig. 16.5B). Precursors of vascular endothelial cells grow into this slit, which ultimately forms the glomerulus. Induced metanephric mesenchyme stimulates the ingrowth of endothelial cells, possibly by the release of a factor similar to FGF. Uninduced mesenchyme does not possess this capability. The endothelial cells are connected with branches from the dorsal aorta, and they form a complex looping structure that ultimately becomes the renal glomerulus. Cells of the glomerular endothelium and the adjoining podocyte epithelium form a thick basement membrane between them. This basement membrane later serves as an important component of the renal filtration apparatus. As the glomerular apparatus of the nephron takes shape, another slit forms in the comma-shaped tubular primordium, thus transforming it into an S-shaped structure (Fig. 16.5C). Cells in the rest of the tubule primordium also undergo an epithelial transformation to form the remainder of the renal tubule. This transformation involves the acquisition of polarity by the differentiating epithelial cells. It is correlated with the deposition of laminin in the extracellular matrix along the basal surface of the cells and the concentration of the integral membrane glycoprotein uvomorulin (E-cadherin), which seals the lateral borders of the cells (Fig. 16.6). As the differentiating tubule assumes an S shape, differing patterns of gene expression are seen along its length. Near the future glomerular end, levels of Pax-2 expression decrease as WT-1 becomes strongly expressed (see Fig. 16.3). Lim-1 expression and the downstream Delta/Notch system are now known to play a prominent role in generating the proximal convoluted tubule. At the other end of the tubule (future distal convoluted tubule), Wnt-4 and E-cadherin remain prominent, whereas in the middle (future proximal convoluted tubule), K-cadherin is a prominent cellular marker. Many of the uninduced mesenchymal cells between tubules undergo apoptosis. Differentiation of the renal tubule progresses from the glomerulus to the proximal and then distal convoluted tubule. During differentiation of the nephron, a portion of the tubule develops into an elongated hairpin loop that extends into the medulla of the kidney as the loop of Henle. As they differentiate, the tubular epithelial cells develop molecular features characteristic of the mature kidney (e.g., brush border antigens or the Tamm-Horsfall glycoprotein [see Fig. 16.4]). Growth of the kidney involves the formation of approximately 15 successive generations of nephrons in its peripheral zone, with the outermost nephrons less mature than the nephrons farther inward. Development of the internal architecture of the kidney is complex, involving the formation of highly ordered arcades of nephrons (Fig. 16.7). Details are beyond the scope of this text. While the many sets of nephrons are differentiating, the kidney becomes progressively larger. The branched system of ducts also becomes much larger and more complex, and it forms the pelvis and system of calyces of the kidney (Fig. 16.8). These structures collect the urine and funnel it into the ureters. During much of the fetal period, the kidneys are divided into grossly visible lobes. By birth, the lobation is already much less evident, and it disappears during the neonatal period. When they first take shape, the metanephric kidneys are located deep in the pelvic region. During the late embryonic and early fetal period, they undergo a pronounced shift in position that moves them into the abdominal region. This shift results partly from actual migration and partly from a marked expansion of the caudal region of the embryo. Two concurrent components to the migration occur. One is a caudocranial shift from the level of L4 to L1 or even the T12 vertebra (Fig. 16.9). The other is a lateral displacement. These changes bring the kidneys into contact with the adrenal glands, which form a cap of glandular tissue on the cranial pole of each kidney. During their migration, the kidneys also undergo a 90-degree rotation, with the pelvis ultimately facing the midline. As they are migrating out of the pelvic cavity, the kidneys slide over the large umbilical arteries, which branch from the caudal end of the aorta. All these changes occur behind the peritoneum because the kidneys are retroperitoneal organs. During the early phases of migration of the metanephric kidneys, the mesonephric kidneys regress. The mesonephric ducts are retained, however, as they become closely associated with the developing gonads. The division of the cloaca into the rectum and urogenital sinus region was introduced in Chapter 15 (see Fig. 15.13). The urogenital sinus is continuous with the allantois, which has an expanded base continuous with the urogenital sinus and an attenuated tubular process that extends into the body stalk on the other end. Along with part of the urogenital sinus, the dilated base of the allantois continues to expand to form the urinary bladder, and its attenuated distal end solidifies into the cordlike urachus, which ultimately forms the median umbilical ligament that leads from the bladder to the umbilical region (see Fig. 16.19). As the bladder grows, its expanding wall, which is derived from tailbud mesenchyme, incorporates the mesonephric ducts and the ureteric buds (Fig. 16.10). The result is that these structures open separately into the posterior wall of the bladder. Through a poorly defined mechanism possibly involving mechanical tension exerted by the migrating kidneys, the ends of the ureters open into the bladder laterally and cephalically to the mesonephric ducts. The region bounded by these structures is called the trigone of the bladder, but much of the substance of the trigone itself is composed of musculature from the bladder. Only small strips of smooth muscle along the edges of the trigone may arise from ureteral smooth muscle. At the entrance of the mesonephric ducts, the bladder becomes sharply attenuated. This region, originally part of the urogenital sinus, forms the urethra, which serves as the outlet of the bladder (see p. 401). Clinical Correlation 16.1 presents congenital anomalies of the urinary system. Clinical Correlation 16.1 Congenital Anomalies of the Urinary System Anomalies of the urinary system are common (3% to 4% of live births). Many are asymptomatic, and others are manifest only later in life. Figure 16.11 summarizes the locations of many frequently encountered malformations of the urinary system. Renal agenesis is the unilateral or bilateral absence of any trace of kidney tissue (Fig. 16.12A). Unilateral renal agenesis is seen in roughly 0.1% of adults, whereas bilateral renal agenesis occurs in 1 in 3000 to 4000 newborns. The ureter may be present. This anomaly is usually ascribed to a faulty inductive interaction between the ureteric bud and the metanephrogenic mesenchyme. As many as 50% of cases of renal agenesis in humans have been attributed to mutations of RET or glial cell line–derived neurotrophic factor (GDNF), which are key players in the earliest induction of the ureteric bud. Individuals with unilateral renal agenesis are often asymptomatic, but typically the single kidney undergoes compensatory hypertrophy to maintain a normal functional mass of renal tissue. An infant born with bilateral renal agenesis dies within a few days after birth. Because of the lack of urine output, reduction in the volume of amniotic fluid (oligohydramnios) during pregnancy is often an associated feature. Infants born with bilateral renal agenesis characteristically exhibit the Potter sequence, consisting of a flattened nose, wide interpupillary space, a receding chin, tapering fingers, low-set ears, hip dislocation, and pulmonary hypoplasia (Fig. 16.13). Respiratory failure from pulmonary hypoplasia is a common cause of neonatal death in this condition, especially when pulmonary hypoplasia is caused by disorders other than renal agenesis. A sequence is classified as a set of malformations secondary to a primary disturbance in development. In the Potter sequence, reduced urinary output secondary to renal agenesis or a urinary blockage is the factor that sets in motion the other disorders seen in this constellation (Fig. 16.14). The actual mechanical effects result from the lack of mechanical buffering by the greatly reduced amount of amniotic fluid. An intermediate condition between renal agenesis and a normal kidney is renal hypoplasia (see Fig. 16.12B), in which one kidney or, more rarely, both kidneys will be substantially smaller than normal even though a certain degree of function may be retained. Although a specific cause for renal hypoplasia has not been identified, some cases may be related to deficiencies in growth factors or their receptors that are active during later critical phases of metanephrogenesis. As with renal agenesis, the normal counterpart to a hypoplastic kidney is likely to undergo compensatory hypertrophy. Renal duplications range from a simple duplication of the renal pelvis to a completely separate supernumerary kidney. Similar to hypoplastic kidneys, renal duplications may be asymptomatic, although the incidence of renal infections may be increased. Many variants of duplications of the ureter have also been described (see Fig. 16.12). Duplication anomalies are commonly attributed to splitting or wide separation of branches of the ureteric bud, the latter resulting from ectopic expression of GDNF more proximally along the mesonephric duct. Anomalies of Renal Migration and Rotation The most common disturbance of renal migration leaves a kidney in the pelvic cavity (Fig. 16.15A). This disturbance is usually associated with malrotation of the kidney, so that the hilus of the pelvic kidney faces anteriorly instead of toward the midline. Another category of migratory malformation is crossed ectopia, in which one kidney and its associated ureter are found on the same side of the body as the other kidney (Fig. 16.15B). In this condition, the ectopic kidney may be fused with the normal kidney. In the condition of horseshoe kidney, which can occur in 1 in 400 individuals, the kidneys are typically fused at their inferior poles (Fig. 16.16). Horseshoe kidneys cannot migrate out of the pelvic cavity because the inferior mesenteric artery, coming off the aorta, blocks them. In most cases, horseshoe kidneys are asymptomatic, but occasionally pain or obstruction of the ureters may occur. This condition may be associated with anomalies of other internal organs. Pelvic kidneys are subject to an increased incidence of infections and obstructions of the ureters. Polycystic Disease of the Kidney Congenital polycystic disease of the kidney occurs in more than 1 in 800 live births and is manifested by the presence of hundreds to thousands of cysts of different sizes within the parenchyma of the kidney (Fig. 16.17). The most common form, autosomal dominant, is the result of mutations of the genes PKD1 and PKD2, which produce the proteins polycystin-1 and polycystin-2. These proteins, which are surface membrane receptors, affect various cellular processes, such as proliferation, polarity, and differentiation. Affected individuals exhibit persisting fetal patterns of location of these proteins, along with receptors for epidermal growth factor and sodium, potassium–adenosine triphosphatase (Na+,K+-ATPase); the result is the budding off of spherical cysts from a variety of locations along the nephron. In some genetic mutants, the cysts are caused by disturbances in the orientation of mitoses within the developing ducts. In normal ductal development, mitoses are aligned along the long axis of the duct. In mutations that result in randomly oriented mitoses, the collecting ducts and even tubules begin to balloon out, forming cysts, instead of elongating. These cysts enlarge and can attain diameters greater than 10 cm. Cysts of other organs, especially the liver and pancreas, are frequently associated with polycystic kidneys. Ureters may open into a variety of ectopic sites (Fig. 16.18). Because of the continuous supply of urine flowing through them, these sites are symptomatic and usually easy to diagnose. Their embryogenesis is commonly attributed to ectopic origins of the ureteric buds in the early embryo. Cysts, Sinuses, and Fistulas of the Urachus If parts of the lumen of the allantois fail to become obliterated, urachal cysts, sinuses, or fistulas can form (Fig. 16.19). In the case of urachal fistula, urine seeps from the umbilicus. Urachal sinuses or cysts may swell in later life if they are not evident in an infant. Exstrophy of the bladder is a major defect in which the urinary bladder opens broadly onto the abdominal wall (Fig. 16.20). Rather than being a primary defect of the urinary system, it is most commonly attributed to an insufficiency of mesodermal tissue of the ventral abdominal wall. Although initially the ventral body wall may be closed with ectoderm, it breaks down in the absence of mesoderm, and degeneration of the anterior wall of the bladder typically follows. In male infants, exstrophy of the bladder commonly involves the phallus, and a condition called epispadias results (see p. 404). A reduction in the expression of sonic hedgehog (shh) signaling in the pericloacal epithelia may contribute to a deficiency of tissue in the bladder and the external genitalia. According to a different hypothesis, aneurysmic swellings of the dorsal aortae in the area may prevent tissues from fusing along the ventral midline and keep the walls of the cloaca from closing around the future bladder. Development of the genital system is one phase in the overall sexual differentiation of an individual (Fig. 16.21). Sexual determination begins at fertilization, when a Y chromosome or an additional X chromosome is joined to the X chromosome already in the egg. This phase represents the genetic determination of gender. Although the genetic gender of the embryo is fixed at fertilization, the gross phenotypic gender of the embryo is not manifested until the seventh week of development. Before that time, the principal morphological indicator of the embryo’s gender is the presence or absence of the sex chromatin (Barr body) in the female. The Barr body is the result of inactivation of one of the X chromosomes. During this morphologically indifferent stage of sexual development, the gametes migrate into the gonadal primordia from the yolk sac. The phenotypic differentiation of gender is traditionally considered to begin with the gonads* and progresses with gonadal influences on the sexual duct systems. Similar influences on the differentiation of the external genitalia and finally on the development of the secondary sexual characteristics (e.g., body configuration, breasts, hair patterns) complete the events that constitute the overall process of sexual differentiation. Sexual differentiation of the brain, which has an influence on behavior, also occurs.
Urogenital System
Urinary System
Early Forms of the Kidney
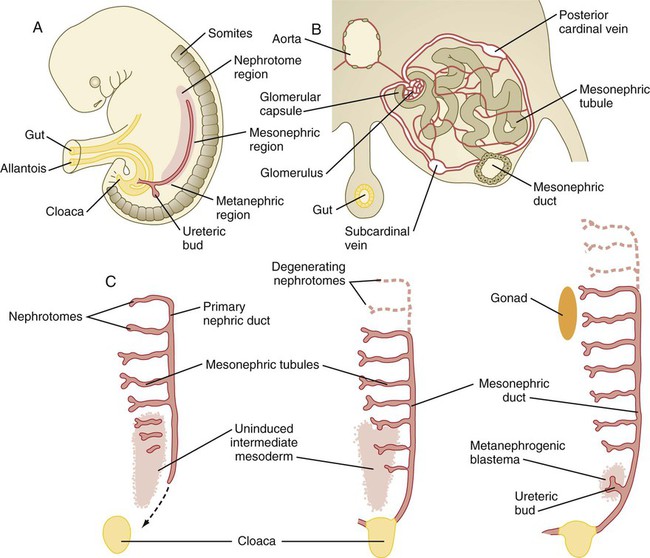
A, Subdivision of the intermediate mesoderm into areas that will form nephrotomes, mesonephros, and metanephros. B, Cross section through mesonephros showing a well-developed mesonephric tubule and its associated vasculature. C, Caudal progression of formation of the mesonephros and degeneration of the most cranial segments of the primitive kidney.
Metanephros
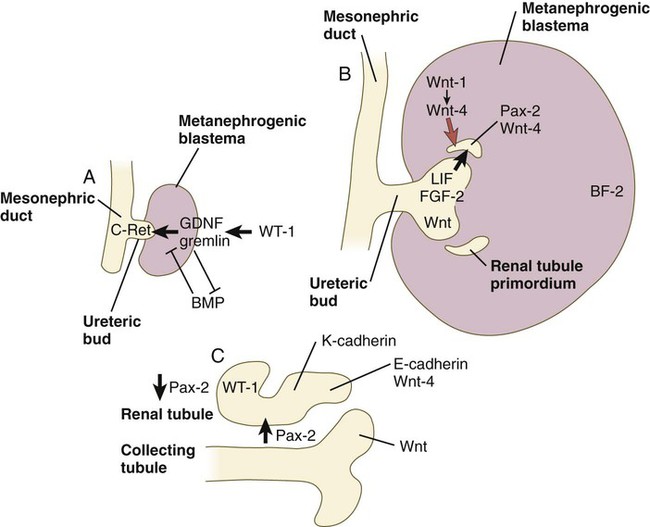
A, Early induction of metanephros. B, Branching of ureteric bud. C, Early tubule formation. FGF, fibroblast growth factor; GDNF, glial cell line–derived neurotrophic factor; LIF, leukemia inhibitory factor.
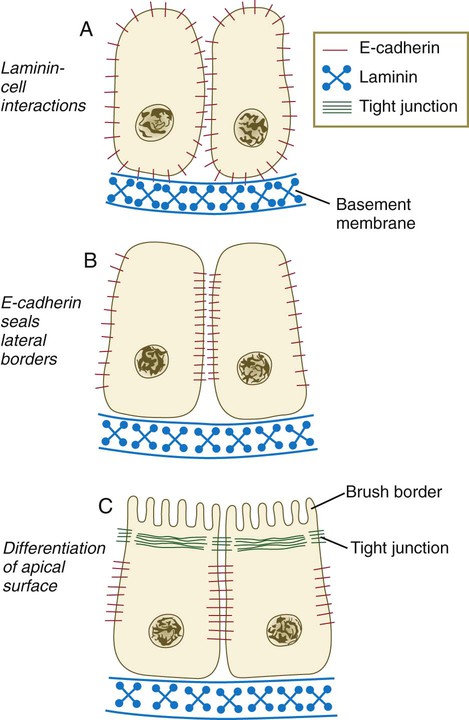
A, Development of polarity is triggered by interactions between laminin and the cell surface, but E-cadherin is still distributed in a nonpolar manner. B, E-cadherin redistribution occurs, and E-cadherin interactions seal the lateral borders of the cells. C, Apical border of epithelial cells differentiates, as seen by formation of a brush border. (Based on Ekblom P: FASEB J 3:2141-2150, 1989.)
Later Changes in Kidney Development
Formation of the Urinary Bladder
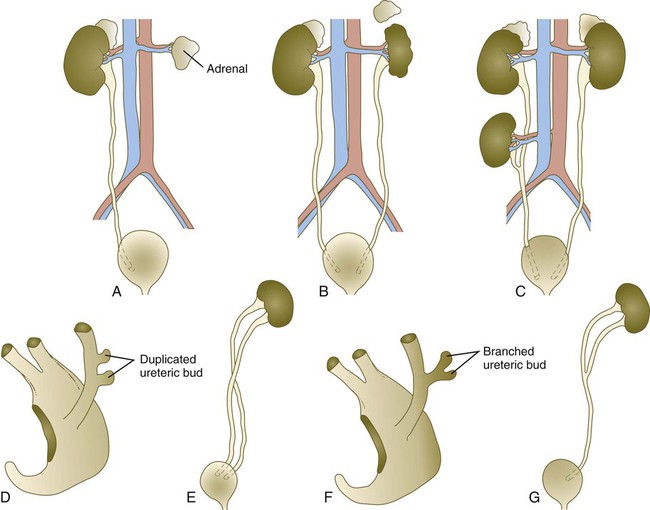
A, Unilateral renal agenesis. The ureter is also missing. B, Unilateral renal hypoplasia. C, Supernumerary kidney. D and E, Complete duplication of ureter, presumably arising from two separate ureteric buds. F and G, Partial duplication of ureter, presumably arising from a bifurcated ureteric bud.
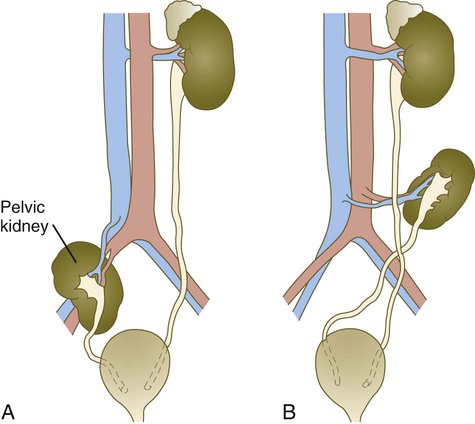
A, Pelvic kidney. B, Crossed ectopia. The right kidney has crossed the left ureter and has migrated only part of the normal distance.
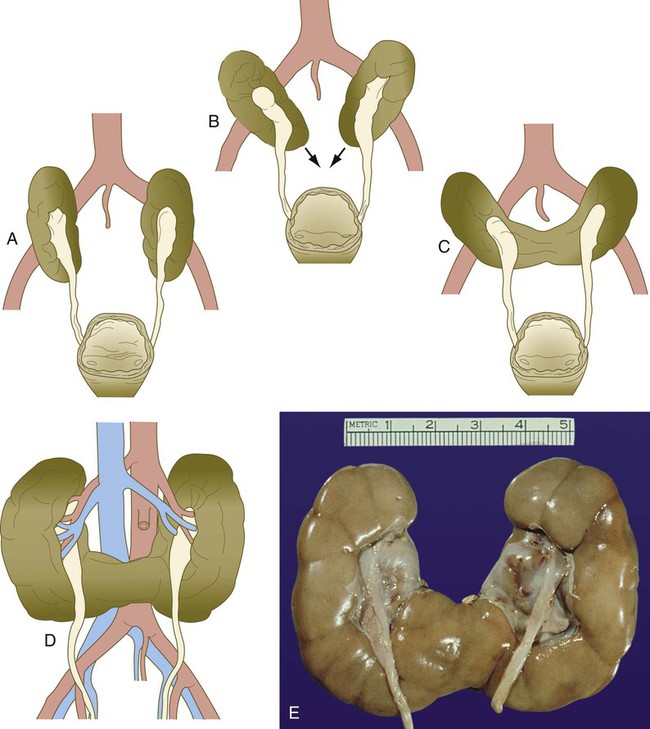
A to C, As the kidneys migrate out of the pelvis, their caudal poles touch and fuse. D, Pelvic kidney in an adult. Note the lack of rotation of the kidneys so that the ureters face ventrally instead of medially. E, Horseshoe kidney. (E, Photo 914E from the Arey-DaPeña Pediatric Pathology Photographic Collection, Human Developmental Anatomy Center, National Museum of Health and Medicine, Armed Forces Institute of Pathology, Washington, D.C.)
Genital System
Urogenital System