Tissues
LANGUAGE OF SCIENCE
 Before reading the chapter, say each of these terms out loud. This will help you avoid stumbling over them as you read.
Before reading the chapter, say each of these terms out loud. This will help you avoid stumbling over them as you read.
[absorp- swallow, -tion process]
[adipo- fat, -ose full of, tissu- fabric]
[alveo- hollow, -ola- little, -ar relating to]
[are- open space, -ola- little, -ar relating to]
[a- without, -vas- vessel, -ula- little, -ar relating to]
[base- base, –ment thing, membran-thin skin]
[canal- channel, -iculi little] pl., canaliculi
[cancel- lattice, -ous characterized by, tissu- fabric]
[cardia- heart, -ac relating to, mus- mouse, -cle small, tissu- fabric]
[chondro- cartilage, -cyte cell]
collagenous dense fibrous tissue
(kah-LAJ-ehnus dense FYE-brus TISH-yoo)
[colla- glue, -gen produce, dense thick, fibro- fiber, -ous relating to, tissu- fabric]
[column- column, -ar relating to]
[compact to put together, tissu- fabric]
[con- together, -nect- bind, -ive relating to, tissu- fabric]
[con- together, -nect- bind, -ive relating to, tissu- fabric, membran- thin skin]
[cub- cube, -oid like, -al relating to]
[cut- skin, -aneous relating to, membran- thin skin].
[dendr- tree, -ite part (branch) of]
[dense thick, fibro- fiber, -ous relating to, connect- bind, -ive relating to, tissu- fabric]
[elast- to drive or beat out, -ic relating to, cartilag cartilage]
(eh-LAS-tik dense FYE-brus TISH-yoo)
[elast- to drive or beat out, -ic relating to, dense thick, fibr fiber, tissu- fabric]
[elast- drive or beat out, -in substance]
(en-doh-KON-dral osi- fi-KAY-shun)
[endo- inward or within, -chondr- cartilage, -al relating to, oss- bone, -fication to make]
[endo- inward or within, -crin-secrete, gland acorn]
[endo- inward or within, -derm skin]
[endo- inward or within, -theli- nipple, -um thing] pl., endothelia
[epi- on or upon, -theli nipple, -al relating to, membran- thin skin]
[epi- on or upon, -theli- nipple, -al relating to, tissu-fabric]
[epi– on or upon, theli- nipple, -um thing] pl., epithelia
[excret- to separate, -tion process]
[exo- outside or outward, -crin- secrete, gland acorn]
(eks-trah-SEL-yoolar MAY-triks)
[extra- beyond, -cell- storeroom, -ular relating to, matrix womb] pl., matrices]
[fibro- fiber, –cartilage cartilage]
[gland- acorn (gland), -ula- little, -ar relating to]
[hema- blood, -poie- to make, -ic relating to, tissu- fabric]
[hist- tissue, -amine ammonia compound]
[histo- tissue, -genesis origin]
[histo- tissue, -o- combining form, –log- words (study of), -y activity]
[hyaline of glass, cartilage cartilage]
[inter- between, -stit- stand, -al relating to]
[lam- plate, -ella little] pl., lamellae
[lamina thin plate, propria proper]
loose fibrous connective tissue
(LOOS FYE-brus koh-NEK-tiv TISH-yoo)
[mast fattening, cell storeroom]
[membran- thin skin, -ous characterized by]
[meso- middle or median, -theli- nipple, -um thing] pl., mesothelia
[micro- small, -villi shaggy hairs] pl., microvilli
[muc- slime, -ous characterized by, membran- thin skin]
[multi- many, -cell- storeroom, -ular relating to, gland acorn]
[mus- mouse, -cle small, tissu- fabric]
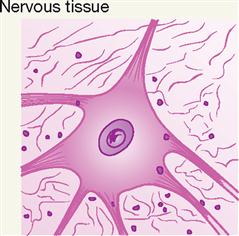
[nervous relating to nerves, tissu- fabric]
[neuro- nerve, -glia glue] sing., neuroglial cell
[peri- around, -chondr- cartilage, -um thing] pl., perichondria
[proteo- protein, -glycan polysaccharide (from -glyc- sweet)]
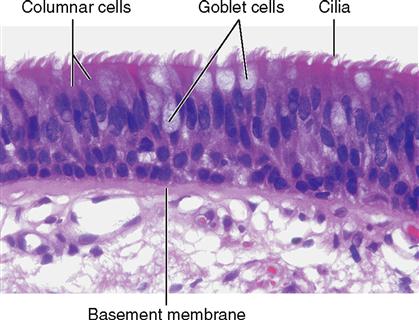
pseudostratified columnar epithelium
(SOOD-oh-STRAT-i-fyed koh-LUM-nar ep-i-THEE-lee-um)
[re- again, generat- produce, -tion process]
[ret- net, -ic– relating to, -ul- little, -ar characterized by, tissu- fabric]
[secret- separate, -tion process]
[sero- watery body fluid, -ous characterized by, membran- thin skin]
[simple not mixed, epi- on, -theli- nipple, -um thing] pl., epithelia
[skeleto- dried body, -al relating to, mus- mouse, -cle small, tissu- fabric]
[mus- mouse, -cle small, tissu- fabric]
[squam- scale, -ous characterized by]
(STRAT-i-fyde ep-i-THEE-lee-um)
[strati- layer, epi- on, -theli- nipple, -um thing] pl., epithelia
[syn- together, -ovi- egg (white), -al relating to, fluid flow]
[syn- together, -ovi- egg (white), -al relating to, membran- thin skin]
[trab– beam, -ula little] pl., trabeculae
(tran-ZISH-en-al ep-i-THEE-lee-um)
[trans- across, -tion process, epi- on, -theli- nipple, -um thing] pl., epithelium
[tub- tube, -ul- little, -ar relating to]
[uni- one, -cell- storeroom, -ular relating to, gland acorn]
LANGUAGE OF MEDICINE
[bio- life, -ops- view, -y act of]
[cancer crab or malignant tumor]
[carcino- cancer, -gen produce]
[chemo- chemical, -therapy treatment]
[hyper- excessive, -plasia shape]
[immuno- free, -therapy treatment]
[laser shortened “light amplification by stimulated emission of radiation”]
[meta- change, -stasis standing]
[peri- around, -ton- to stretch, -itis inflammation]
[pleur- rib, -itis inflammation]
(ray-dee-AY-shun THAYR-ah-pee)
[radiat- to emit rays, -tion process, therapy treatment]
[tumor swelling, suppress- press down, -or agent, gen- produce or generate]
INTRODUCTION TO TISSUES
Each tissue specializes in performing at least one unique function that helps maintain homeostasis, ensuring the survival of the whole body. The arrangement of cells in one tissue may form a thin sheet only one cell deep, whereas the cells of another tissue may form huge masses containing millions of cells. Regardless of the size, shape, or arrangement of cells in a tissue, they all are surrounded by or embedded in a complex extracellular material that often is called simply matrix.
The four major types of human tissue that were introduced in Chapter 1 are described in more detail in this chapter. An understanding of the major tissue types will help you understand the next higher levels of organization in the body—organs and organ systems. Eventually, your knowledge of histology (the biology of tissues) will give you a better appreciation for the nature of the whole body.
This brief introduction to histology involves many images made with microscopes of various types. Refer to Tools of Microscopic Anatomy online at A&P Connect for an overview of the types of microscopy used in this chapter.

PRINCIPAL TYPES OF TISSUE
Although a number of subtypes are present in the body, all tissues can be classified by their structure and function into four principal types:
The major tissue types are also summarized in Table 6-1.
TABLE 6-1
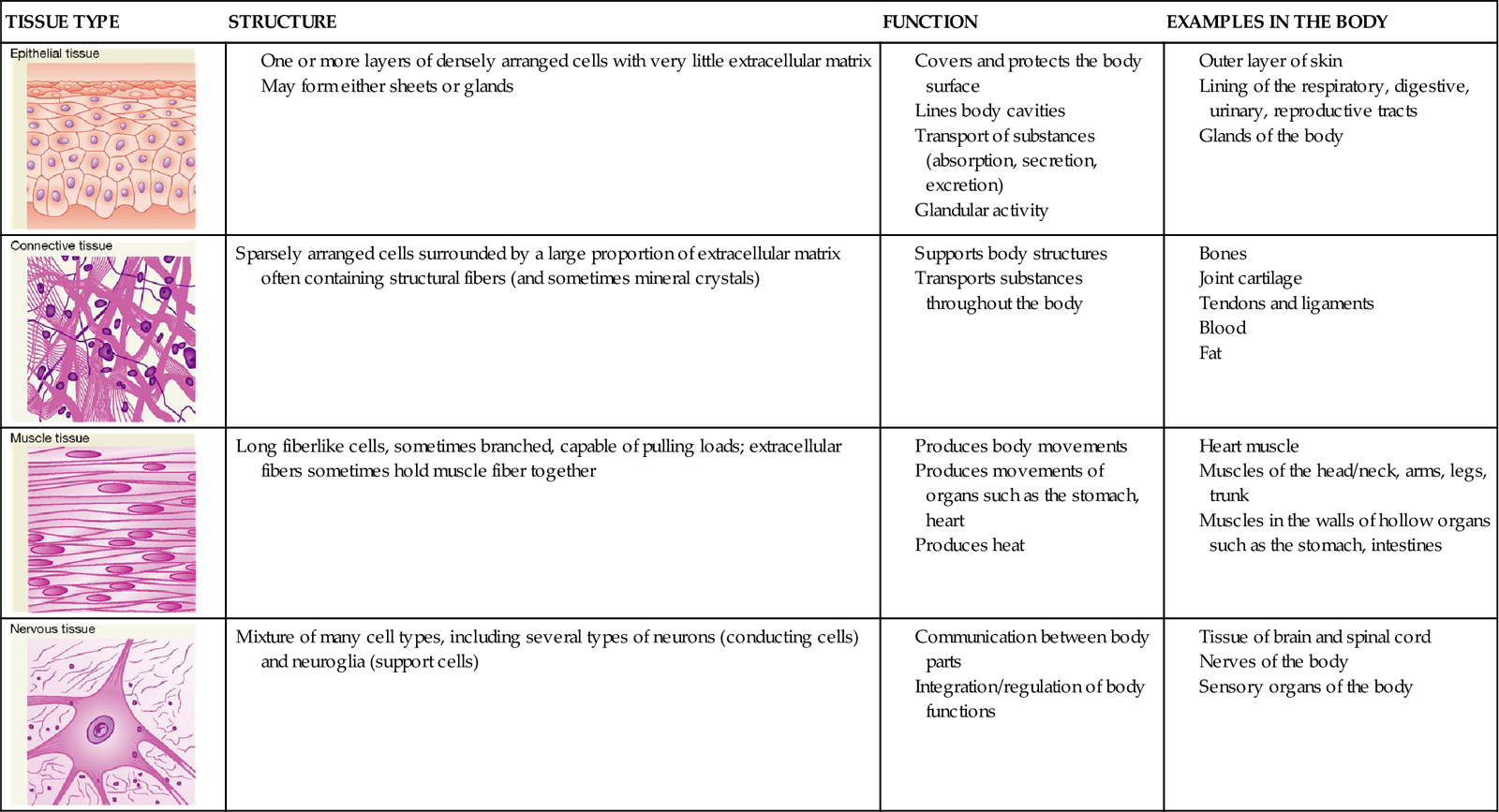
The four major tissues of the body appear early in the embryonic period of development. Within the first 2 weeks after conception, cells of the offspring move and regroup in an orderly way into three primary germ layers called endoderm, mesoderm, and ectoderm. During this process the cells in each germ layer become increasingly more differentiated to form specific tissues—a process called histogenesis (Box 6-1). Remodeling themselves by a combination of new growth, differentiation, and apoptosis, these early layers eventually give rise to the various organs of the body.
Embryonic development is discussed more thoroughly in Chapter 36. For now, take a look at a diagram illustrating the concept of primary germ layers in Embryonic Development of Tissues online at A&P Connect.

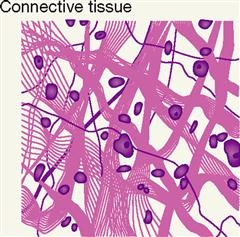
EXTRACELLULAR MATRIX
Tissues differ in the amount and kind of material between the cells—the extracellular matrix (ECM). Figure 6-1 hints at the complex nature of the ECM. The particular makeup of the extracellular matrix in bone and cartilage, for example, contributes to the strength and resiliency of the body. Some tissues have very little ECM. Other tissues are almost entirely extracellular matrix—with only a few cells present. Some types of ECM contain large numbers of structural protein fibers that make them flexible or elastic, some contain many mineral crystals that make them rigid, and others are very fluid.
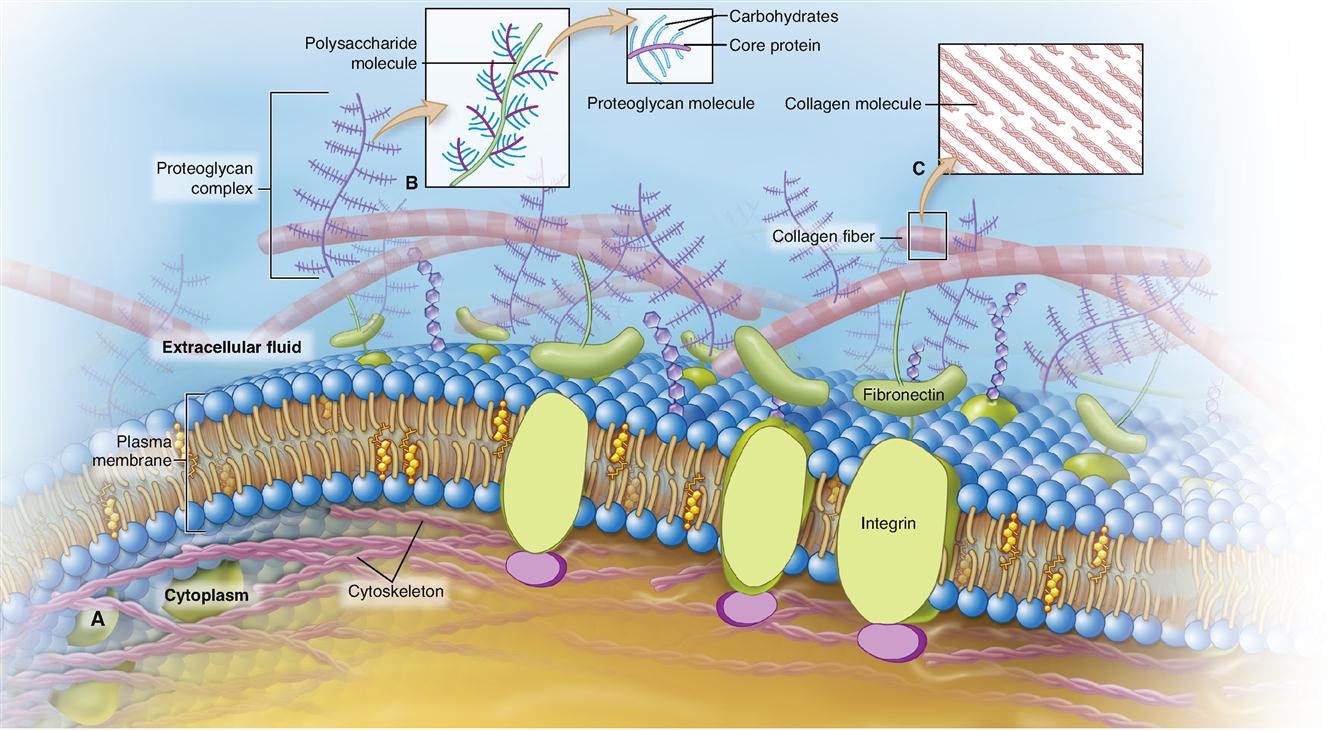
It is only recently that scientists have discovered the complexities of the ECM and its important role in human biology. Besides water, the ECM is made up mostly of proteins and proteoglycans (Table 6-2).
TABLE 6-2
Components of the Extracellular Matrix (ECM)*
| COMPONENT | EXAMPLE | DESCRIPTION | FUNCTION | EXAMPLE OF LOCATION |
| Water | Water molecules along with a small number of ions (mostly Na+ and Cl−) | Solvent for dissolved ECM components; provides fluidity of ECM | All tissues of the body | |
| Proteins and glycoproteins | Collagen | Strong, flexible structural protein fiber | Provides flexible strength to tissues | Tendons, ligaments, bones, cartilage, many tissues |
| Elastin | Flexible, elastic structural protein fiber | Allows flexibility and elastic recoil of tissues | Skin, cartilage of ear, walls of arteries | |
| Fibronectin | Rodlike glycoprotein | Binds ECM to cells; communicates with cells through integrins | Many tissues of the body, for example, connective tissues | |
| Laminin | Glycoproteins arranged as a three-pronged fork | Binds ECM components together and to cells; communicates with cells through integrins | In basal lamina (basement membrane) of epithelial tissues | |
| Proteoglycans | Various types | Protein backbone with attached chains of various polysaccharides: | ||
| Chondroitin sulfate | Shock absorber | Cartilage, bone, heart valves | ||
| Heparin | Reduces blood clotting | Lining of some arteries | ||
| Hyaluronate | Thickens fluid; lubricates | Loose fibrous connective tissue, joint fluids |
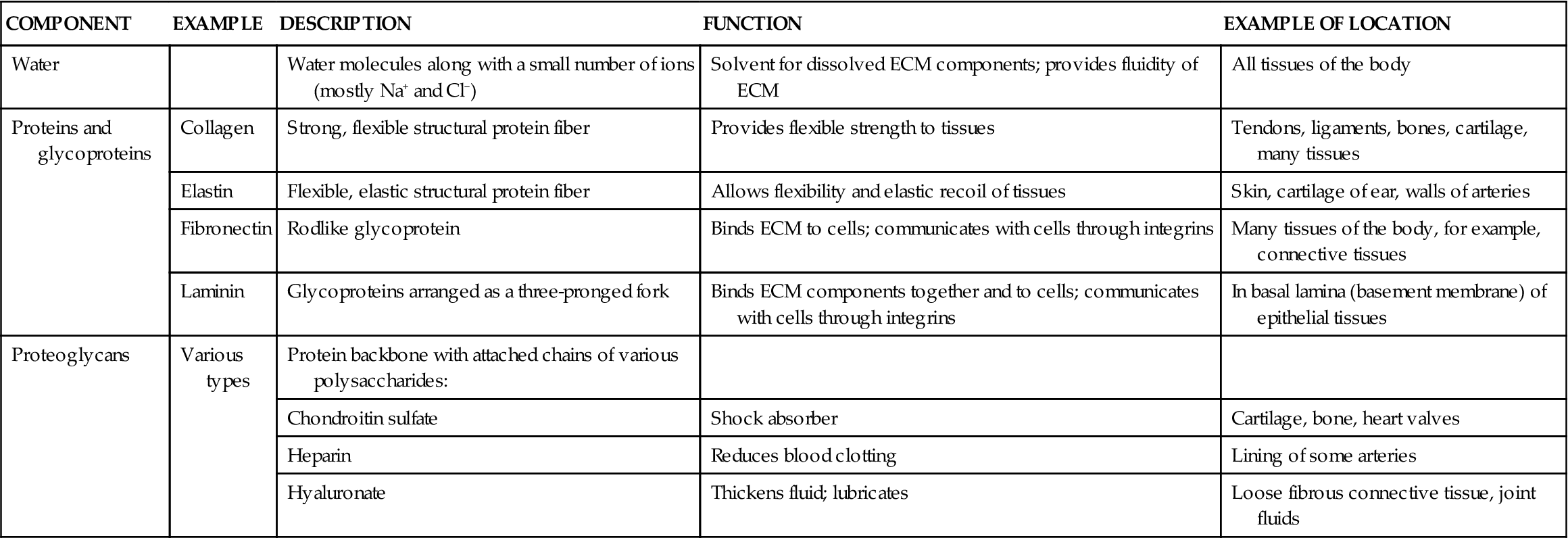
*Not all components are present in all ECM; examples of only some of the many major ECM components are provided.
Proteins in the extracellular matrix include various types of structural protein fibers such as collagen and elastin, both of which are discussed frequently throughout this book. The ECM also contains many glycoproteins, which are mainly protein molecules with attached carbohydrate subunits. For example, fibronectin and laminin both help connect collagen and proteoglycans to cells by attaching to integrin molecules embedded in the plasma membrane of cells (see Figure 6-1). This arrangement thus unites the cells and their surroundings into an integral structure. It also provides a communication mechanism between the ECM and the cell that allows coordination of cell and tissue development, as well as guidance of cell movement and shape changes.
Proteoglycans are hybrid molecules made up mostly of carbohydrates attached to a protein backbone, as you can see in Figure 6-1, B. Many of the sugars attached to the protein backbone of a proteoglycan molecule are N-acetylglucosamine (NAG). Examples of proteoglycans are chondroitin sulfate, heparin, and hyaluronate (see Table 6-2).
In some tissues, such as bone, there may also be calcium-containing mineral crystals that make the ECM rigid. Considering the makeup of ECM, it’s no wonder that many dietary supplements that claim to improve the health of bones and joints include calcium, glucosamine, and/or chondroitin sulfate—all are important components of the ECM of cartilage and bone tissues.
In some tissues, it is the ECM that holds the tissue in a single mass. For example, in skeletal muscles, it is mostly a network of structural protein fibers in the ECM that holds skeletal muscle tissue together. In such cases, components of the ECM bind to the integrins in the outer membranes of the cells, and the integrins bind to components of the internal cytoskeleton.
In other tissues, it is primarily the intercellular junctions, such as the desmosomes and tight junctions described in a previous chapter (see Chapter 3, pp. 83–84), that hold groups of cells together to form tissues found in sheets or other continuous masses of cells. The tissue that forms the outer layer of skin is held together this way. In some tissues, the ECM does not bind to tissue cells. For example, the fluid nature of the blood’s matrix (plasma) does not hold blood tissue in a solid mass at all.
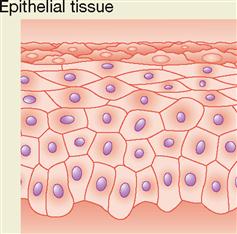
EPITHELIAL TISSUE
Types and Locations of Epithelial Tissue
Epithelial tissue, or epithelium, often is subdivided into two types: (1) membranous (covering or lining) epithelium and (2) glandular epithelium. Membranous epithelium covers the body and some of its parts and lines the serous cavities (pleural, pericardial, and peritoneal), the blood and lymphatic vessels, and the respiratory, digestive, and genitourinary tracts. Glandular epithelium is grouped in solid cords or hollow follicles that form the secretory units of endocrine and exocrine glands.
Functions of Epithelial Tissue
Epithelial tissues have a widespread distribution throughout the body and serve several important functions:
Generalizations About Epithelial Tissue
Most epithelial tissues are characterized by extremely limited amounts of intercellular, or matrix, material. This explains their characteristic appearance, when viewed under a light microscope, of a continuous sheet of cells packed tightly together. With the electron microscope, however, narrow spaces—about 20 nanometers (one millionth of an inch) wide—can be seen around the cells. These spaces, like other intercellular spaces, contain interstitial fluid (IF).
Epithelial tissues generally renew themselves throughout life. The presence of stem cells in most epithelial tissues allows epithelium to continuously produce new cells of various types.
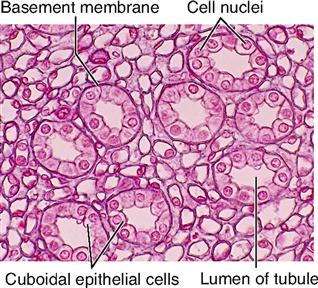
Sheets of epithelial cells compose the surface layer of skin and mucous and serous membranes. The epithelial tissue attaches to an underlying layer of connective tissue by means of a thin noncellular layer of adhesive, permeable material called the basement membrane (BM) (Figure 6-2). Both epithelial and connective tissue cells synthesize the basement membrane, which is a highly complex structure made up partly of glycoprotein material secreted by the epithelial components and a fine mesh of fibers produced by the connective tissue cells. Histologists refer to the glycoprotein material secreted by epithelial cells as the basal lamina and to the connective tissue fibers as the fibroreticular lamina. The union of basal and fibroreticular lamina forms the basement membrane. Integrins embedded in each plasma membrane help bind the cytoskeletons of the epithelial cells to the fibers of the basement membrane so that a strong connection forms between them.
Epithelial tissues contain no blood vessels. As a result, epithelium is said to be avascular (a, “without”; vascular, “vessels”). Hence oxygen and nutrients must diffuse from capillaries in the underlying connective tissue through the permeable basement membrane to reach living epithelial cells.
At intervals between adjacent epithelial cells, their plasma membranes are modified to hold the cells together. These complex intercellular structures, such as desmosomes and tight junctions, are described in Chapter 3. Epithelial cells can reproduce themselves. They frequently go through the process of cell division. Because epithelial cells in many locations meet considerable wear and tear, this fact has practical importance. It means, for example, that new cells can replace old or destroyed epithelial cells in the skin or in the lining of the gut or respiratory tract.
Cross sections of epithelial tissues are described in Box 6-2.
Box 6-2
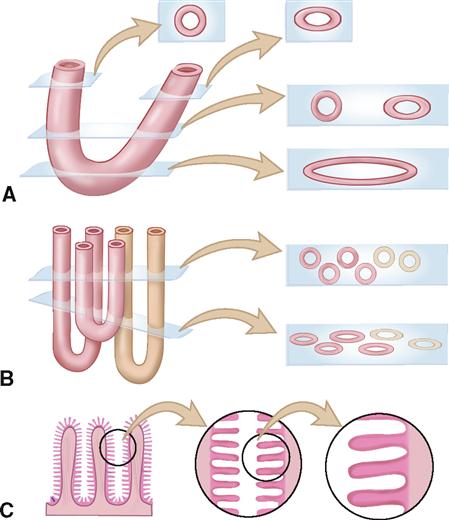
When you look at photomicrographs of epithelial tissues or other structures that are tubelike, saclike, or folded into complex shapes, it is sometimes hard to imagine what you are really looking at. As diagram A shows, when you cut a tube on a cross section, the slice looks like a ring (if cut at a right angle) or oval (if cut at an oblique, or slanted, angle). Diagram A also shows that if the tube is bent where the cut is made, it can also look like an oval on your slide. Diagram B shows what happens when you have many tubes next to one another—your slice has many round or oval rings (depending on the angle of the cut). Compare Diagram A to Figure 6-5. Can you imagine the type of tissue that produced this slice? Diagram C shows what can happen when a membrane such as the intestinal lining is folded into complex shapes. The slice may look like a sort of zigzag line of cells, or if it is at a high magnification, it may just look like a series of parallel rows. Look at Figure 6-6. Can you imagine what the original tissue must have looked like? Look ahead to Figures 28-9 through 28-21. Can you “see” the context or “situation” of the various tissue slices found in many of these figures? Try sketching out the “big picture” of the context of various tissue samples shown in the photomicrographs of this chapter, and then keep them in your notebook. By doing so, you’ll be preparing yourself for later chapters (and later courses) by learning to identify the context of a tissue “on sight.”
Classification of Epithelial Tissue
MEMBRANOUS EPITHELIUM
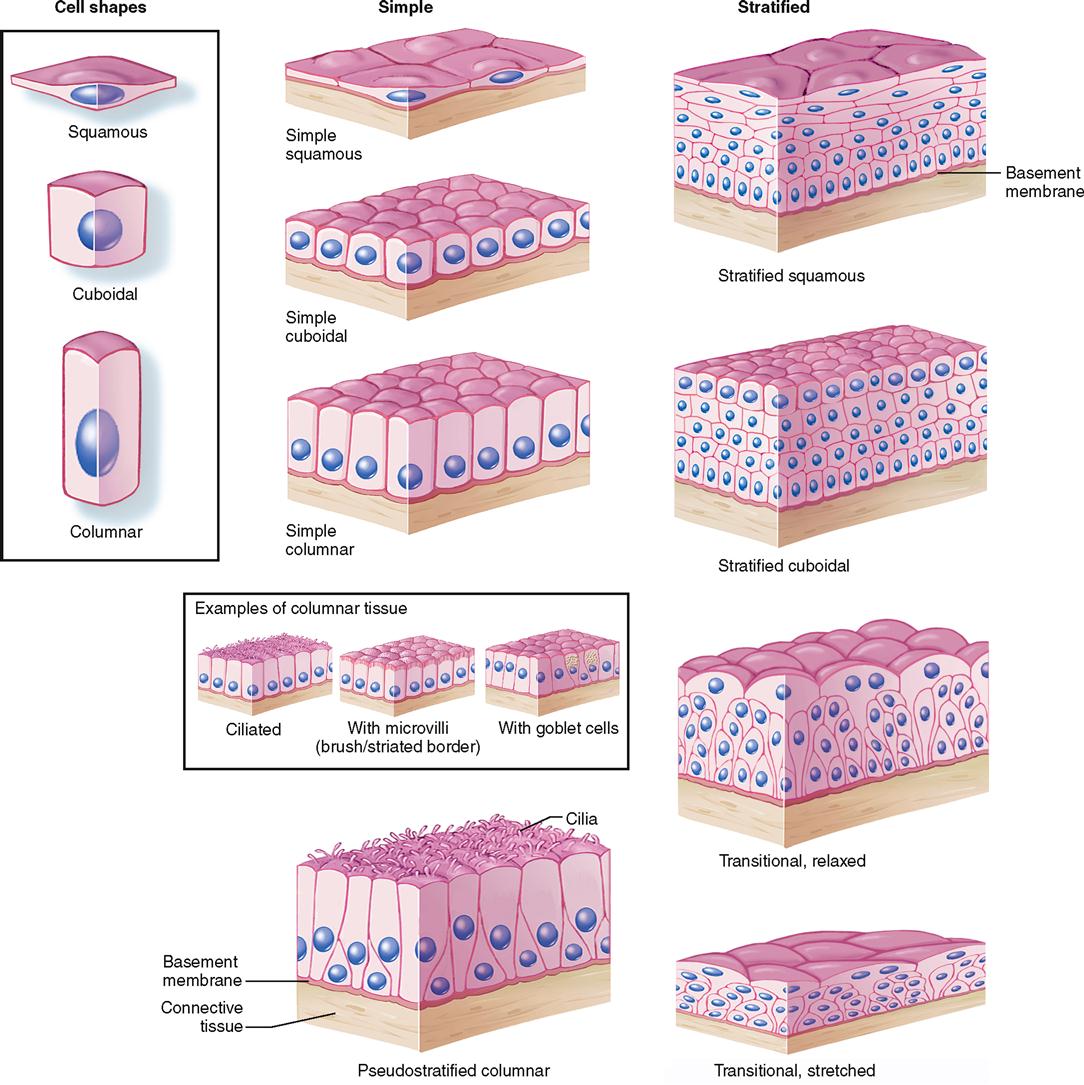
Classification Based on Cell Shape
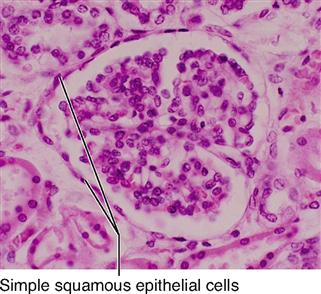
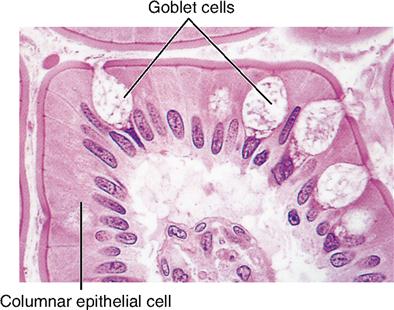
The shape of membranous epithelial cells may be used for classification purposes. Four cell shapes, called squamous, cuboidal, columnar, and pseudostratified columnar, are used in this classification scheme (see Figure 6-2). Squamous (Latin, “scaly”) cells are flat and platelike. Cuboidal cells, as the name implies, are cube-shaped and have more cytoplasm than the scalelike squamous cells do. Columnar epithelial cells have more height than width and thus appear narrow and cylindrical. Pseudostratified columnar epithelium has only one layer of oddly shaped columnar cells. Although each cell touches the basement membrane, the tops of some pseudostratified cells do not fully extend to the surface of the membrane. Also, some nuclei are near the “top” of the cell and some near the “bottom” of the cell—rather than all nuclei being near the bottom. The result is a false (pseudo) appearance of layering, or stratification, when only a single layer of cells is present.
Classification Based on Layers of Cells
In most cases the location and function of membranous epithelium determine whether its cells will be stacked and layered or arranged in a sheet one cell layer thick. An arrangement of epithelial cells in a single layer is called simple epithelium. If epithelial cells are layered one on another, the tissue is called stratified epithelium. Transitional epithelium (described later) is a unique arrangement of differing cell shapes in a stratified, or layered, epithelial sheet.
If membranous or covering epithelium is classified by the shape and layering of its cells, the specific types listed in Table 6-3 are possible. Notice that stratified tissue types are named for the shape of cells in their top layer only. Each type is described in the paragraphs that follow, and selected examples are illustrated in Figures 6-3 to 6-10.
TABLE 6-3
Classification Scheme for Membranous Epithelial Tissues
| SHAPE OF CELLS* | TISSUE TYPE |
| One Layer | |
| Squamous | Simple squamous |
| Cuboidal | Simple cuboidal |
| Columnar | Simple columnar |
| Pseudostratified columnar | Pseudostratified columnar |
| Several Layers | |
| Squamous | Stratified squamous |
| Cuboidal | Stratified cuboidal |
| Columnar | Stratified columnar |
| (Varies) | Transitional |
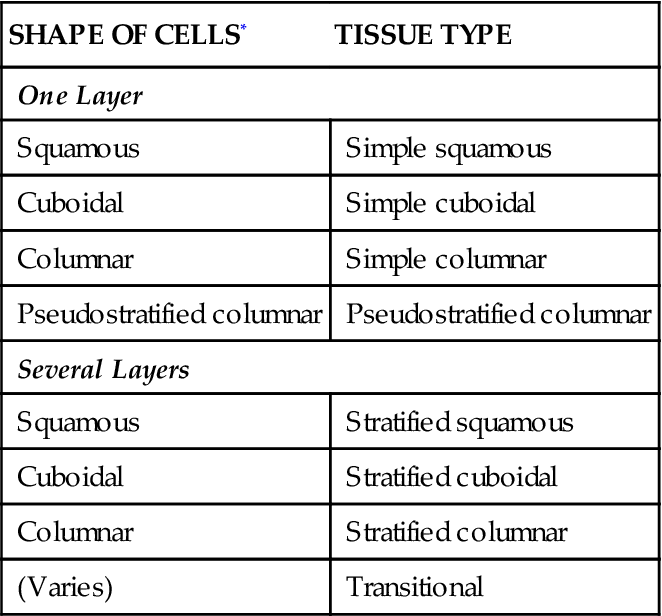
*In the top layer (if more than one layer is present in the tissue).