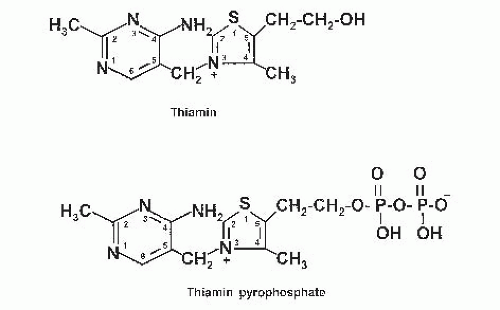
Certain foods contain thiaminases—thermolabile enzymes that rapidly degrade thiamin (
3). Thiaminase I is encountered in some raw fish, shellfish, and ferns, as well as in microorganisms such as
Clostridium thiaminolyticus. Thiaminase II, which has an action distinct from that of thiaminase I, is found in other organisms such as
Candida aneurinolytica. Thiaminases act during food storage or during passage through the gastrointestinal tract. Consequently, regular consumption of raw fish (with or without fermentation), raw shellfish, and ferns is a risk factor for the development of thiamin deficiency. Antithiamin compounds are thermostable and have been identified in some ferns, teas, and betel nut, in which the toxic agents were found to be analogs of polyphenolic compounds such as tannic acid (tannin).