CHAPTER 21 The Male Reproductive System
The Clonal Nature of Male Germ Cells
The male reproductive system consists of the testes, genital ducts, accessory glands, and penis (Figure 21–1). Testes produce sperm but also contain endocrine cells secreting hormones such as testosterone, which drives male reproductive physiology. Testosterone is important for spermatogenesis, sexual differentiation during embryonic and fetal development, and control of gonadotropin secretion in the pituitary. A metabolite of testosterone, dihydrotestosterone, also begins to act on many tissues during puberty (eg, male accessory glands and hair follicles).
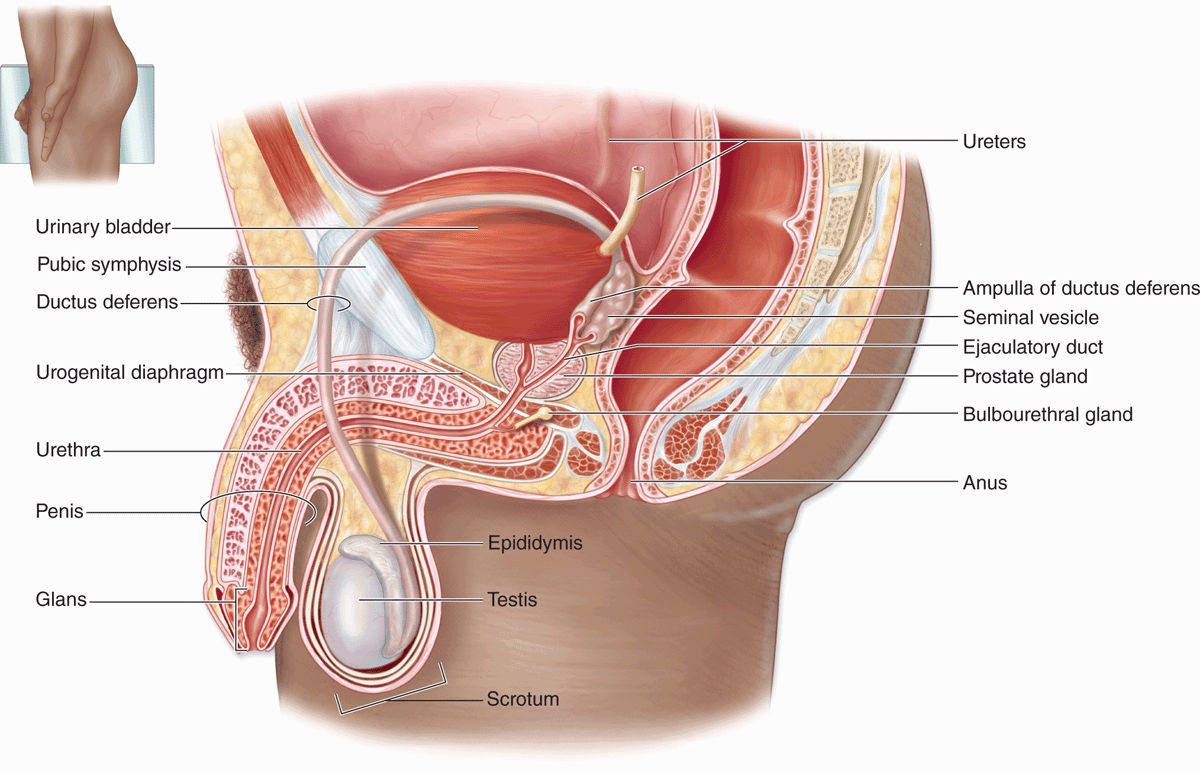
FIGURE 21–1 The male reproductive system.
The genital ducts and accessory glands produce secretions required for sperm activity and contract to propel spermatozoa and the secretions from the penile urethra. These secretions provide nutrients for spermatozoa while they are confined to the male reproductive tract. Spermatozoa and the secretions of the accessory glands make up the semen (L, seed), which is introduced into the female reproductive tract by the penis.
TESTES
Each testis (or testicle) is surrounded by a dense connective tissue capsule, the tunica albuginea, which thickens on the posterior side to form the mediastinum testis. From this fibrous region, septa penetrate the organ and divide it into about 250 pyramidal compartments or testicular lobules (Figures 21–2 and 21–3). Each lobule contains sparse connective tissue with endocrine interstitial cells (or Leydig cells) secreting testosterone, and one to four highly convoluted seminiferous tubules in which sperm production occurs.
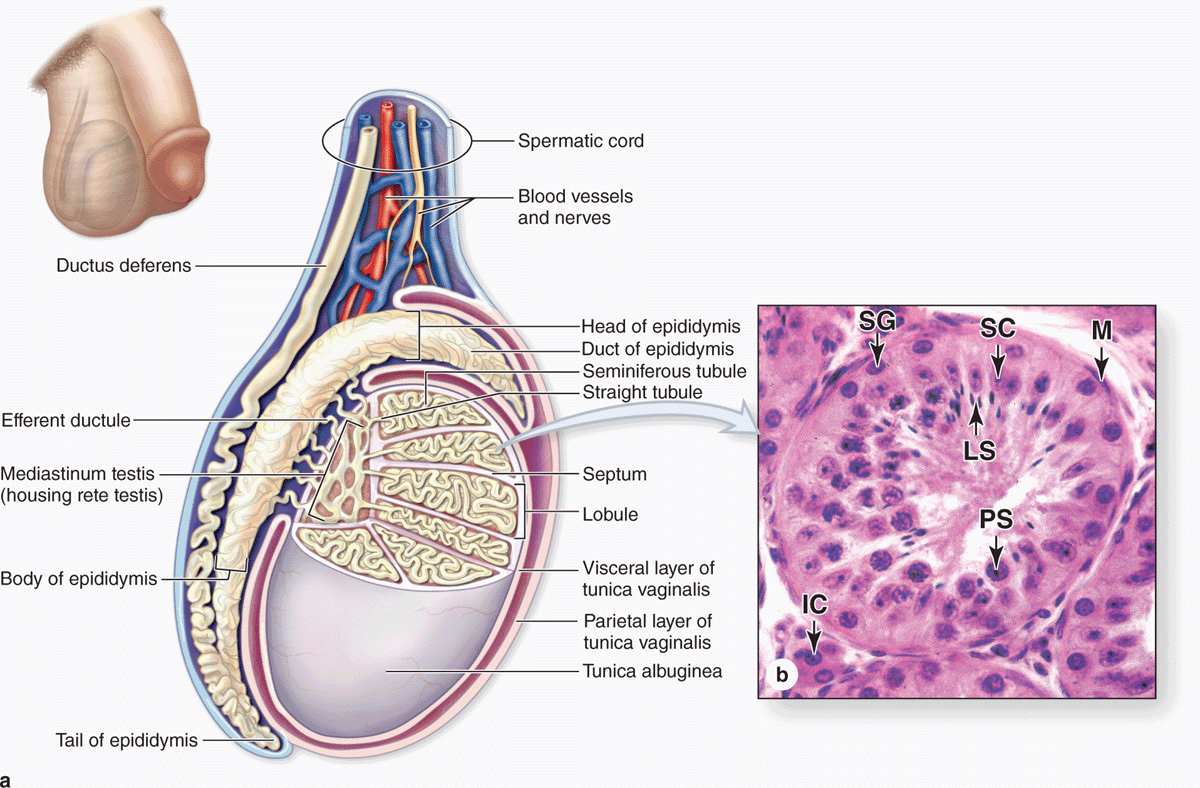
FIGURE 21–2 Testes and seminiferous tubules.
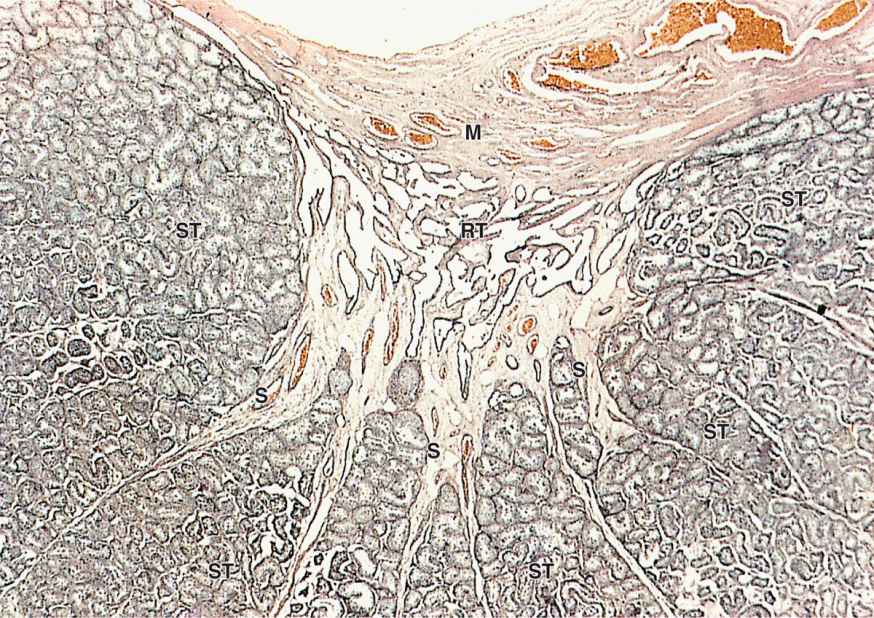
FIGURE 21–3 Lobules converging at rete testis.
The testes develop retroperitoneally in the dorsal wall of the embryonic abdominal cavity and are moved during fetal development to become suspended in the two halves of the scrotal sac, or scrotum, at the ends of the spermatic cords (Figure 21–2). During migration from the abdominal cavity, each testis carries with it a serous sac, the tunica vaginalis, derived from the peritoneum. This tunic consists of an outer parietal layer lining the scrotum and an inner visceral layer, covering the tunica albuginea on the anterior and lateral sides of the testis (Figure 21–2).
Having evolved in cold-blooded animals, certain molecular events in the process of sperm formation cannot occur at the core body temperature of 37°C. A permissive temperature of about 34°C is maintained in the scrotal sac by various mechanisms. Each testicular artery is surrounded by a rich pampiniform venous plexus containing cooler blood from the testis, which draws heat from the arterial blood by a countercurrent heat-exchange system. Evaporation of sweat from the scrotum also contributes to heat loss. Relaxation or contraction of the thin dartos muscle of the scrotum and the cremaster muscles of the spermatic cords move the testes away from or closer to the body, respectively, allowing further control on testicular temperature.
 MEDICAL APPLICATION
MEDICAL APPLICATION
An excessive accumulation of serous fluid in one or both sides of the scrotal sac, termed a hydrocele, is the most common cause of scrotal swelling and a condition easily corrected surgically.
Cryptorchidism (Gr. kryptos, hidden + orchis, testis), the failure of one or both testes to descend from the abdomen, occurs in about 4% of male neonates, but in most of these individuals the testes move to the scrotum during the first year. Bilateral cryptorchidism causes infertility if not surgically corrected by 2 to 3 years of age.
Interstitial Tissue
The interstitial tissue of the testis between the seminiferous tubules consists of sparse connective tissue containing fibroblasts, lymphatics, and blood vessels including fenestrated capillaries. During puberty interstitial cells, or Leydig cells, develop as large round or polygonal cells with central nuclei and eosinophilic cytoplasm rich in small lipid droplets (Figures 21–2b and 21–4. These cells produce the steroid hormone testosterone, which promotes development of the secondary male sex characteristics. Testosterone is synthesized by enzymes present in the smooth ER and mitochondria similar to the system in adrenal cortical cells.
Testosterone secretion by interstitial cells is triggered by the pituitary gonadotropin, luteinizing hormone (LH), which is also called interstitial cell stimulating hormone (ICSH). Testosterone synthesis thus begins at puberty, when the hypothalamus begins producing gonadotropin-releasing hormone. In the late embryonic testes gonadotropin from the placenta stimulates interstitial cells to synthesize the testosterone needed for development of the ducts and glands of the male reproductive system. These fetal interstitial cells are very active during the third and fourth months of pregnancy, then regress and become quiescent cells resembling fibroblasts until puberty when they resume testosterone synthesis in response to the pituitary gonadotropin.
 MEDICAL APPLICATION
MEDICAL APPLICATION
Both interstitial cell tumors and Sertoli cell tumors are rare. Most (95%) testicular cancer involves germ cell tumors, which only appear after puberty and are much more likely to develop in men with untreated cryptorchidism.
Seminiferous Tubules
Sperm are produced in the seminiferous tubules at a rate of about 2 × 108 per day in the young adult. Each testis has from 250 to 1000 such tubules in its lobules, and each tubule measures 150 to 250 μm in diameter and 30 to 70 cm in length. The combined length of the tubules of one testis totals about 250 m. Each tubule is actually a loop linked by a very short, narrower segment, the straight tubule, to the rete testis, a labyrinth of epithelium-lined channels embedded in the mediastinum testis (see Figures 21–2a and 21–3). About 10-20 efferent ductules connect the rete testis to the head of the epididymis (Figure 21–2a).
Each seminiferous tubule is lined with a complex, specialized stratified epithelium called germinal or spermatogenic epithelium (Figure 21–2b). The basement membrane of this epithelium is covered by fibrous connective tissue, with an innermost layer containing flattened, smooth muscle-like myoid cells (Figure 21–2b), which allow weak contractions of the tubule. The germinal epithelium consists of two types of cells:
 Large nondividing Sertoli cells (Figure 21–4c, d), which physically and metabolically support developing sperm cell precursors.
Large nondividing Sertoli cells (Figure 21–4c, d), which physically and metabolically support developing sperm cell precursors.
 Dividing cells of the spermatogenic lineage (Figure 21–5a).
Dividing cells of the spermatogenic lineage (Figure 21–5a).
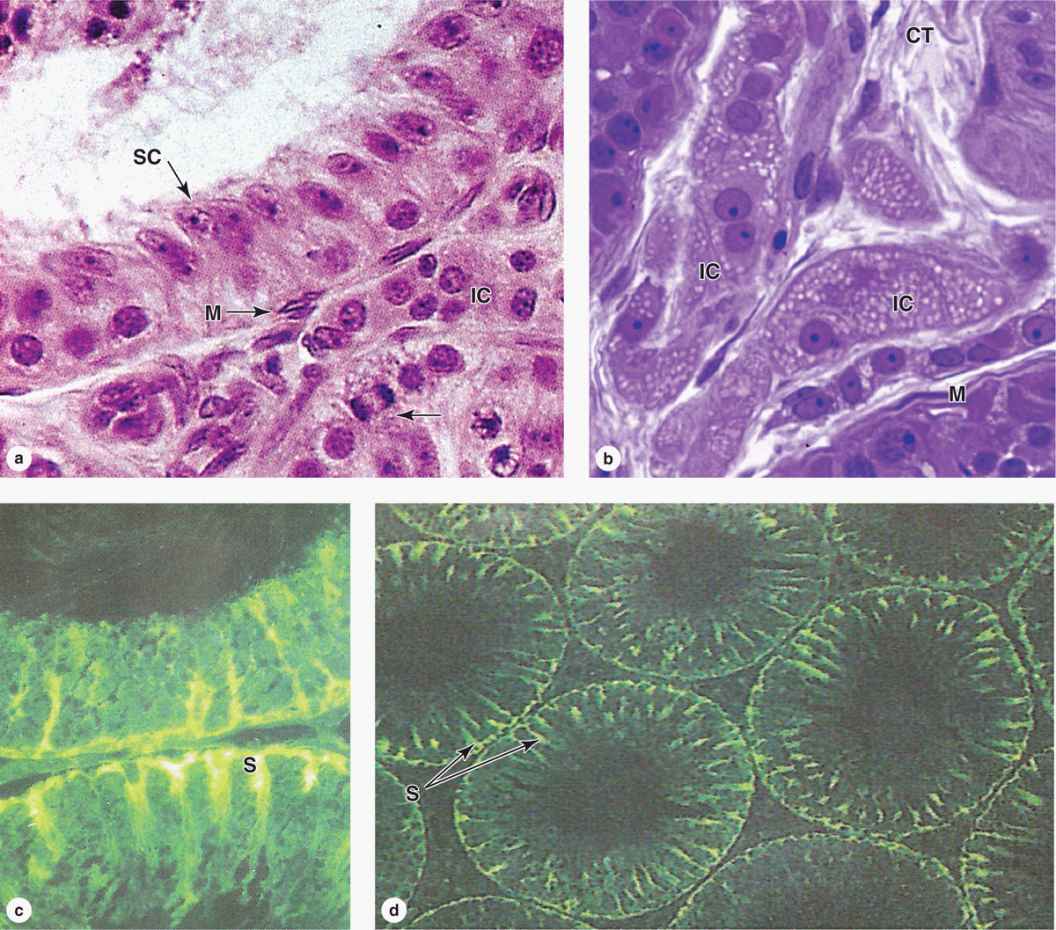
FIGURE 21–4 Seminiferous tubule and interstitial cells.
The cells of the spermatogenic lineage comprise four to eight concentric cell layers and produce the cells that become sperm. As shown in Figure 21–5, spermatogenesis is the first part of sperm production, including stem cell mitosis and meiosis, and spermiogenesis is the final differentiation process occurring in the haploid male germ cells.
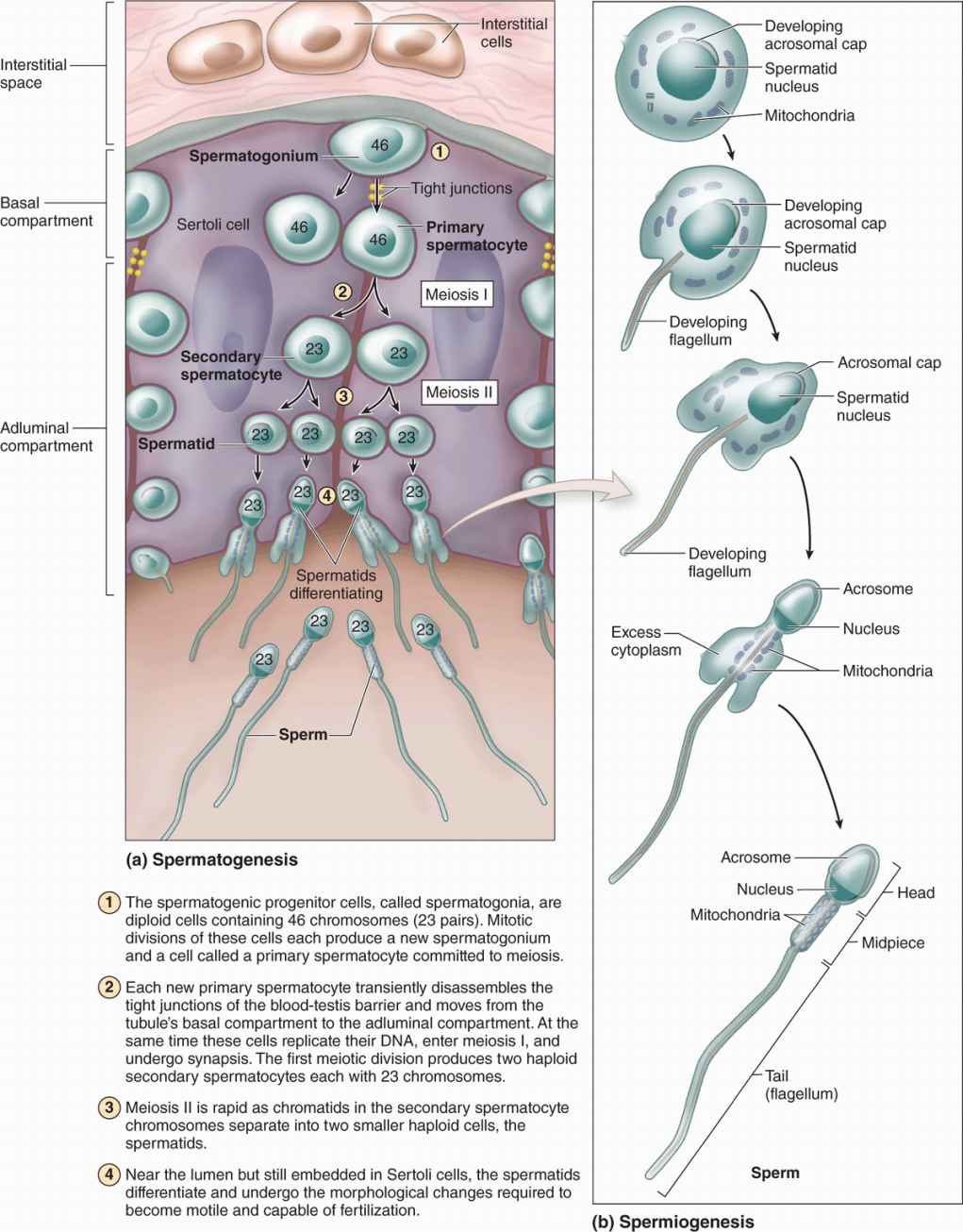
FIGURE 21–5 Spermatogenesis and spermiogenesis.
Spermatogenesis
Spermatogenesis begins at puberty with proliferation of stem and progenitor cells called spermatogonia (Gr. sperma + gone, generation), small round cells about 12 μm in diameter. These cells occupy a basal niche in the epithelial wall of the tubules, next to the basement membrane and closely associated with Sertoli cell surfaces (Figures 21–5, 21–6and 21–7).
Different stages of spermatogonia development can be recognized by subtle changes in shape and staining properties of their nuclei. Spermatogonia with dark, ovoid nuclei act as stem cells, dividing infrequently and giving rise both to new stem cells and to cells with more pale-staining, ovoid nuclei that divide more rapidly as transit amplifying (progenitor) cells (Figure 21–7). These type A spermatogonia each undergo several unique clonal divisions that leave most of the cells interconnected as a syncytium. These become type B spermatogonia, which have more spherical and pale nuclei.
Each type B spermatogonium then undergoes a final mitotic division to produce two cells that grow in size and become primary spermatocytes, which are spherical cells with euchromatic nuclei (Figures 21–6 and 21–7). Primary spermatocytes replicate their DNA, so each chromosome consists of duplicate chromatids, and enter meiosis, during which homologous chromosomes come together in synapsis, DNA recombination occurs, and two rapid cell divisions produce haploid cells (see Chapter 3).
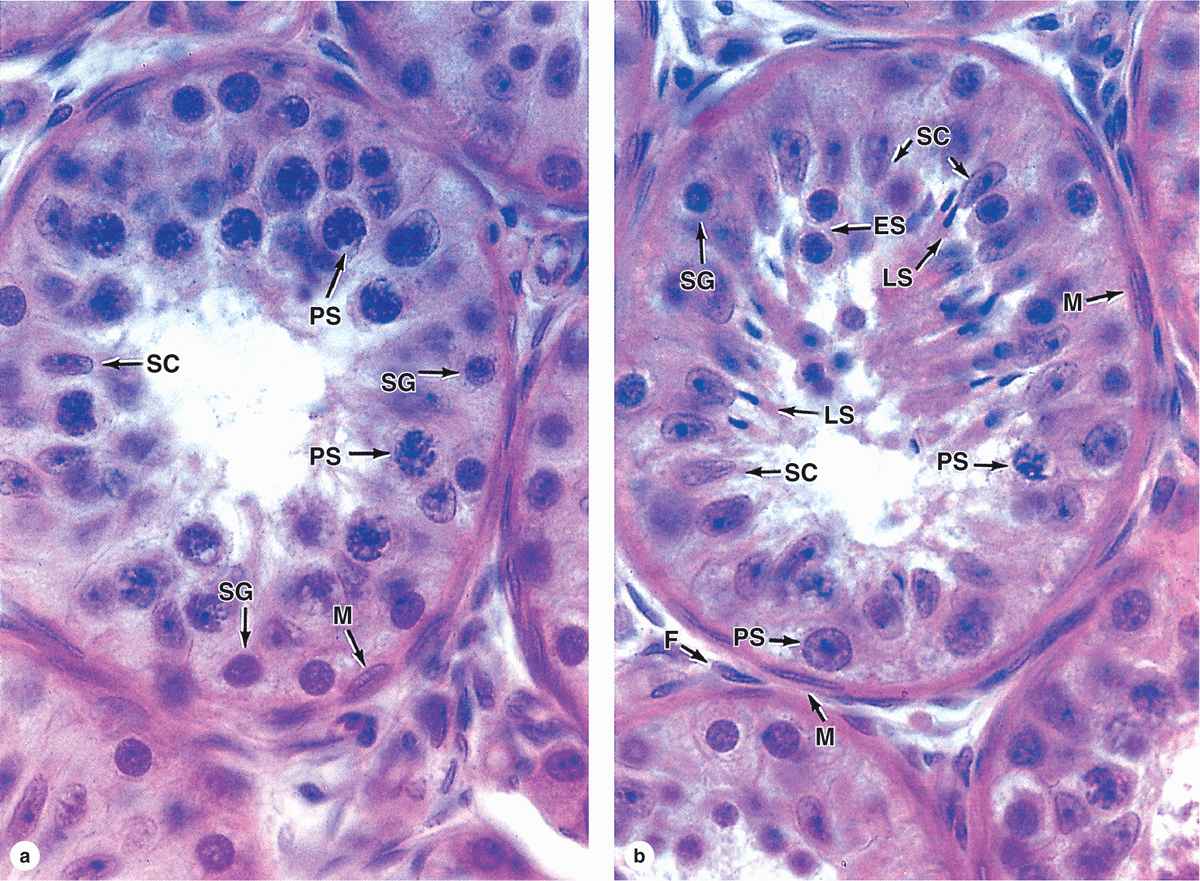
FIGURE 21–6 Seminiferous tubules: Sertoli cells and spermatogenesis.
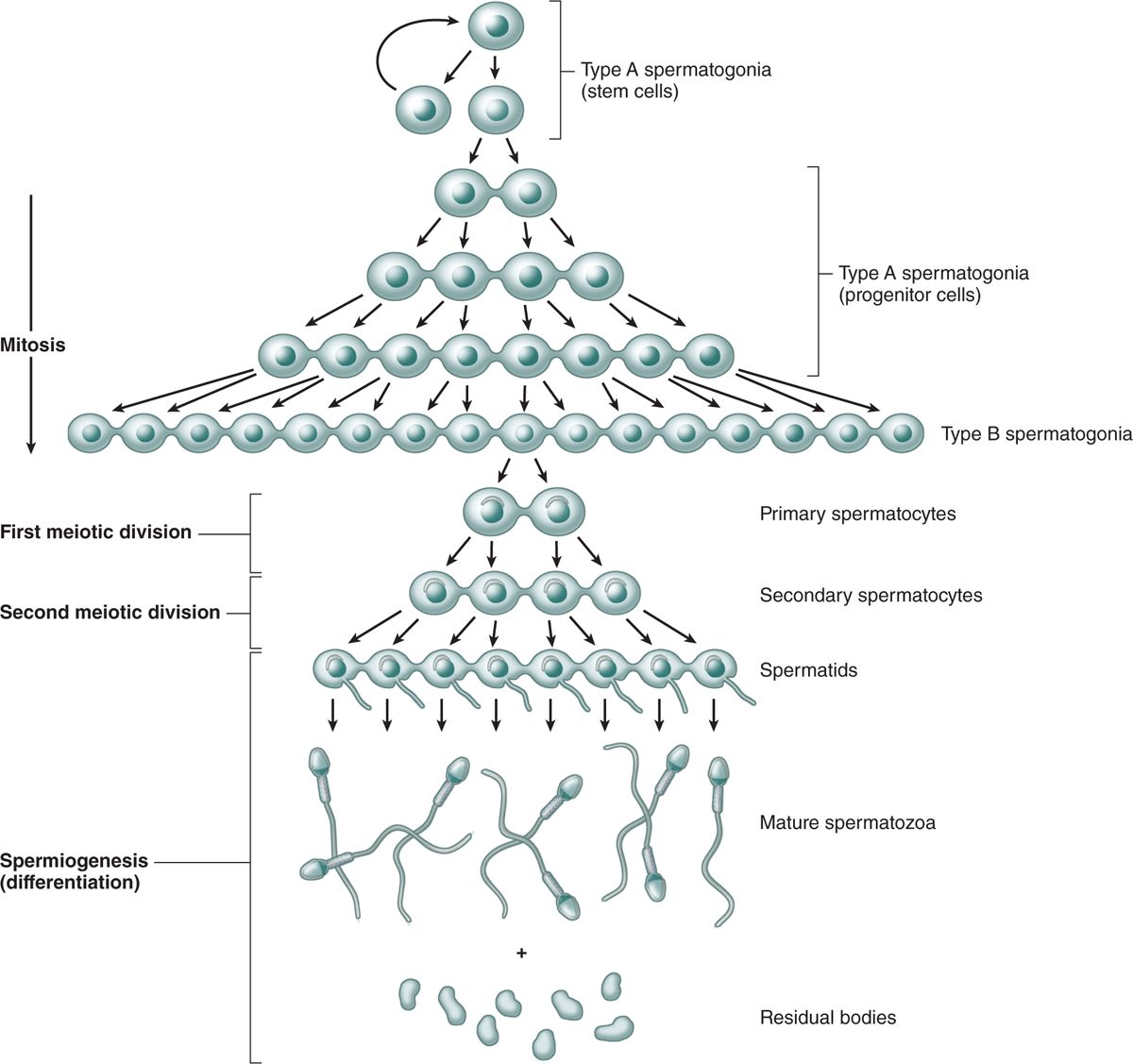
FIGURE 21–7 Clonal nature of spermatogenesis.
The primary spermatocyte has 46 (44 + XY) chromosomes, the diploid number, and a DNA content of 4N. (The letter N denotes either the haploid number of chromosomes, 23 in humans, or the amount of DNA in this set.) Soon after their formation, these cells enter the first meiotic prophase that lasts about 3 weeks. Most spermatocytes seen in sections of testis are in this phase of meiosis. The primary spermatocytes are the largest cells of the spermatogenic lineage and are characterized by the presence of partially condensed chromosomes in various stages of synapsis and recombination (Figure 21–6).
Homologous chromosomes separate in the first meiotic division, which produces smaller cells called secondary spermatocytes (Figures 21–5a and 21–7) with only 23 chromosomes (22 + X or 22 + Y), but each still consists of two chromatids so the amount of DNA is 2N (see Chapter 3). Secondary spermatocytes are rare in testis sections because they are very short-lived cells that remain in interphase only briefly and quickly undergo the second meiotic division.
Division of each secondary spermatocyte separates the chromatids of each chromosome and produces two haploid cells called spermatids each of which contains 23 chromosomes (Figures 21–5a, 21-6, and 21–7). Because no S phase (DNA replication) occurs between the first and second meiotic divisions, the amount of DNA per cell is reduced by half when the chromatids separate and the cells formed are haploid (1N). With fertilization, a haploid ovum and sperm produced by meiosis unite and the normal diploid chromosome number is restored.
The Clonal Nature of Male Germ Cells
The stem cells produced by mitotic divisions of spermatogonia remain as separate cells. However, all subsequent divisions of the daughter cells that become transit amplifying progenitor cells have incomplete cytokinesis after telophase and the cells remain attached to one another by intercellular bridges of cytoplasm (Figure 21–7). These allow free cytoplasmic communication among the cells during their remaining mitotic and meiotic divisions. Although some cells degenerate without completing spermatogenesis and some cells may separate, clones of approximately a hundred cells may remain linked through meiosis. The complete significance of this spermatogenic syncytium is not clear, but the cytoplasmic bridges allow the haploid cells to be supplied with products of the complete diploid genome, including proteins and RNA encoded by genes on the X or Y chromosome missing in their haploid nuclei. The germ cells finally become separated from one another during differentiation (Figure 21–7).
The cellular events and changes between the final mitoses of spermatogonia and the formation of spermatids take about 2 months. The spermatogenic cells are not randomly distributed in the spermatogenic epithelium. Cells at different stages of development are typically grouped together along the tubule, with the intercellular bridges helping to coordinate their divisions and differentiation.
Spermiogenesis
Spermiogenesis, the final phase of sperm production, is the temperature-sensitive process by which spermatids differentiate into spermatozoa, which are highly specialized to deliver male DNA to the ovum. No cell division occurs during this process, and as with spermatogenesis the cells involved remain associated with Sertoli cells.
The haploid spermatids are small (7-8 μm in diameter) cells near the lumen of the seminiferous tubules (Figures 21–5a and 21-6b). Spermiogenesis includes formation of the acrosome (Gr. akron, extremity + soma, body), condensation and elongation of the nucleus, development of the flagellum (L, whip), and the loss of much of the cytoplasm. The end result is the mature spermatozoon, which is released from the Sertoli cell surface into the tubule’s lumen. Spermiogenesis is commonly divided into four phases:
 In the Golgi phase the cytoplasm contains a prominent Golgi apparatus near the nucleus, mitochondria, paired centrioles, and free ribosomes. Small proacrosomal vesicles from the Golgi apparatus coalesce as a single membrane-limited acrosomal cap close to one end of the nucleus (Figures 21–5b and 21–8). The centrioles migrate to a position farthest from the acrosomal cap and one acts as a basal body, organizing the axoneme of the flagellum which is structurally and functionally similar to that of a cilium (see Chapter 2).
In the Golgi phase the cytoplasm contains a prominent Golgi apparatus near the nucleus, mitochondria, paired centrioles, and free ribosomes. Small proacrosomal vesicles from the Golgi apparatus coalesce as a single membrane-limited acrosomal cap close to one end of the nucleus (Figures 21–5b and 21–8). The centrioles migrate to a position farthest from the acrosomal cap and one acts as a basal body, organizing the axoneme of the flagellum which is structurally and functionally similar to that of a cilium (see Chapter 2).
 In the cap phase the acrosomal cap spreads over about half of the condensing nucleus (Figures 21–5b and 21–8). The acrosome is a specialized type of lysosome containing hydrolytic enzymes, mainly hyaluronidase and a trypsin-like protease called acrosin. These enzymes are released when a spermatozoon encounters an oocyte and the acrosomal membrane fuses with the sperm’s plasma membrane. They dissociate cells of the corona radiata and digest the zona pellucida, both structures that surround the egg (see Chapter 22). This process, the acrosomal reaction, is one of the first steps in fertilization.
In the cap phase the acrosomal cap spreads over about half of the condensing nucleus (Figures 21–5b and 21–8). The acrosome is a specialized type of lysosome containing hydrolytic enzymes, mainly hyaluronidase and a trypsin-like protease called acrosin. These enzymes are released when a spermatozoon encounters an oocyte and the acrosomal membrane fuses with the sperm’s plasma membrane. They dissociate cells of the corona radiata and digest the zona pellucida, both structures that surround the egg (see Chapter 22). This process, the acrosomal reaction, is one of the first steps in fertilization.
 In the acrosome phase the head of the developing sperm, containing the acrosome and the condensing nucleus, remains embedded in the Sertoli cell while the growing axoneme extends into the lumen of the tubule (Figure 21–6b). Nuclei become more elongated and very highly condensed, with the histones of nucleosomes replaced by small basic peptides called protamines. Flagellum growth continues as the tail and mitochondria aggregate around its proximal region to form a thickened middle piece where the ATP for flagellar movements is generated (Figure 21–5).
In the acrosome phase the head of the developing sperm, containing the acrosome and the condensing nucleus, remains embedded in the Sertoli cell while the growing axoneme extends into the lumen of the tubule (Figure 21–6b). Nuclei become more elongated and very highly condensed, with the histones of nucleosomes replaced by small basic peptides called protamines. Flagellum growth continues as the tail and mitochondria aggregate around its proximal region to form a thickened middle piece where the ATP for flagellar movements is generated (Figure 21–5).
 In the maturation phase of spermiogenesis, unneeded cytoplasm is shed as a residual body from each spermatozoon and remaining intercellular bridges are lost. Mature but not yet functional sperm (Figure 21–5) are released into the lumen of the tubule.
In the maturation phase of spermiogenesis, unneeded cytoplasm is shed as a residual body from each spermatozoon and remaining intercellular bridges are lost. Mature but not yet functional sperm (Figure 21–5) are released into the lumen of the tubule.
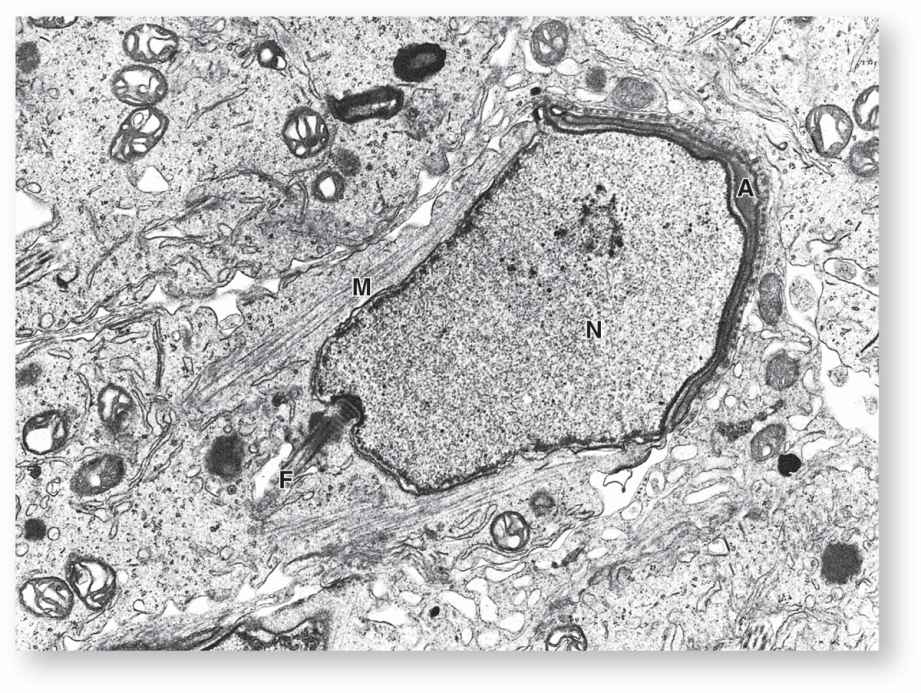
FIGURE 21–8 Differentiating spermatid.



