The skin is the largest single organ of the body, typically accounting for 15% to 20% of total body weight and, in adults, presenting 1.5 to 2 m2 of surface to the external environment. Also known as the integument (L. integumentum, covering) or cutaneous layer, the skin is composed of the epidermis, an epithelial layer of ectodermal origin, and the dermis, a layer of mesodermal connective tissue (Figure 18–1). At the irregular junction between the dermis and epidermis, projections called dermal papillae interdigitate with invaginating epidermal ridges to strengthen adhesion of the two layers. Epidermal derivatives include hairs, nails, and sebaceous and sweat glands. Beneath the dermis lies the subcutaneous tissue or hypodermis (Gr. hypo, under + derma, skin), a loose connective tissue layer usually containing pads of adipocytes. The subcutaneous tissue binds the skin loosely to the underlying tissues and corresponds to the superficial fascia of gross anatomy.
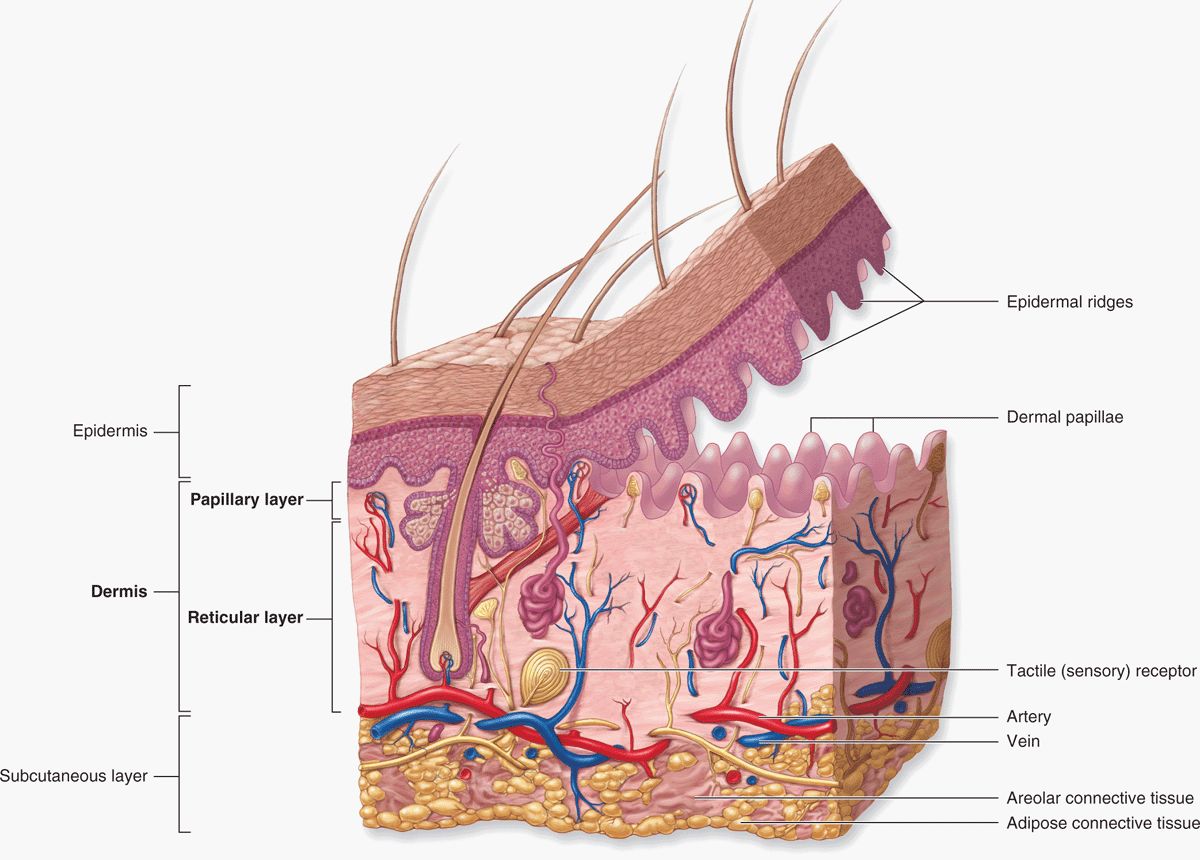
FIGURE 18–1 Layers and appendages of skin.
The specific functions of the skin fall into several broad categories.
 Protective: It provides a physical barrier against thermal and mechanical insults such as friction and against most potential pathogens and other material. Microorganisms that do penetrate skin alert resident lymphocytes and antigen-presenting cells (APCs) in skin and an immune response is mounted. The dark pigment melanin in the epidermis protects cell nuclei from ultraviolet (UV) radiation. Skin is also a permeability barrier against excessive loss or uptake of water, which has allowed for terrestrial life. Skin’s selective permeability allows some lipophilic drugs such as certain steroid hormones and medications to be administered via skin patches.
Protective: It provides a physical barrier against thermal and mechanical insults such as friction and against most potential pathogens and other material. Microorganisms that do penetrate skin alert resident lymphocytes and antigen-presenting cells (APCs) in skin and an immune response is mounted. The dark pigment melanin in the epidermis protects cell nuclei from ultraviolet (UV) radiation. Skin is also a permeability barrier against excessive loss or uptake of water, which has allowed for terrestrial life. Skin’s selective permeability allows some lipophilic drugs such as certain steroid hormones and medications to be administered via skin patches.
 Sensory: Many types of sensory receptors allow skin to constantly monitor the environment, and various skin mechanoreceptors help regulate the body’s interactions with physical objects.
Sensory: Many types of sensory receptors allow skin to constantly monitor the environment, and various skin mechanoreceptors help regulate the body’s interactions with physical objects.
 Thermoregulatory: A constant body temperature is normally easily maintained thanks to the skin’s insulating components (eg, the fatty layer and hair on the head) and its mechanisms for accelerating heat loss (sweat production and a dense superficial microvasculature).
Thermoregulatory: A constant body temperature is normally easily maintained thanks to the skin’s insulating components (eg, the fatty layer and hair on the head) and its mechanisms for accelerating heat loss (sweat production and a dense superficial microvasculature).
 Metabolic: Cells of skin synthesize vitamin D3, needed in calcium metabolism and proper bone formation, through the local action of UV light on the vitamin’s precursor. Excess electrolytes can be removed in sweat, and the subcutaneous layer stores a significant amount of energy in the form of fat.
Metabolic: Cells of skin synthesize vitamin D3, needed in calcium metabolism and proper bone formation, through the local action of UV light on the vitamin’s precursor. Excess electrolytes can be removed in sweat, and the subcutaneous layer stores a significant amount of energy in the form of fat.
 Sexual signaling: Many features of skin, such as pigmentation and hair, are visual indicators of health involved in attraction between the sexes in all vertebrate species, including humans. The effects of sex pheromones produced by the apocrine sweat glands and other skin glands are also important for this attraction.
Sexual signaling: Many features of skin, such as pigmentation and hair, are visual indicators of health involved in attraction between the sexes in all vertebrate species, including humans. The effects of sex pheromones produced by the apocrine sweat glands and other skin glands are also important for this attraction.
The dermal-epidermal interdigitations are of the peg-and-socket variety in most skin (Figure 18–1), but they occur as well-formed ridges and grooves in the thick skin of the palms and soles, which is more subject to friction. These ridges and the intervening sulci form distinctive patterns unique for each individual, appearing as combinations of loops, arches, and whorls, called dermatoglyphs, also known as fingerprints and footprints. Skin is elastic and can expand rapidly to cover swollen areas and, like the gut lining, is self-renewing throughout life. In healthy individuals injured skin is repaired rapidly. The molecular basis of skin healing is increasingly well understood and provides a basis for better understanding of repair and regeneration in other organs.
EPIDERMIS
The epidermis consists mainly of a stratified squamous keratinized epithelium composed of cells called keratinocytes. There are also three much less abundant epidermal cell types: pigment-producing melanocytes, antigen-presenting Langerhans cells, and tactile epithelial cells called Merkel cells (Figure 18–2).
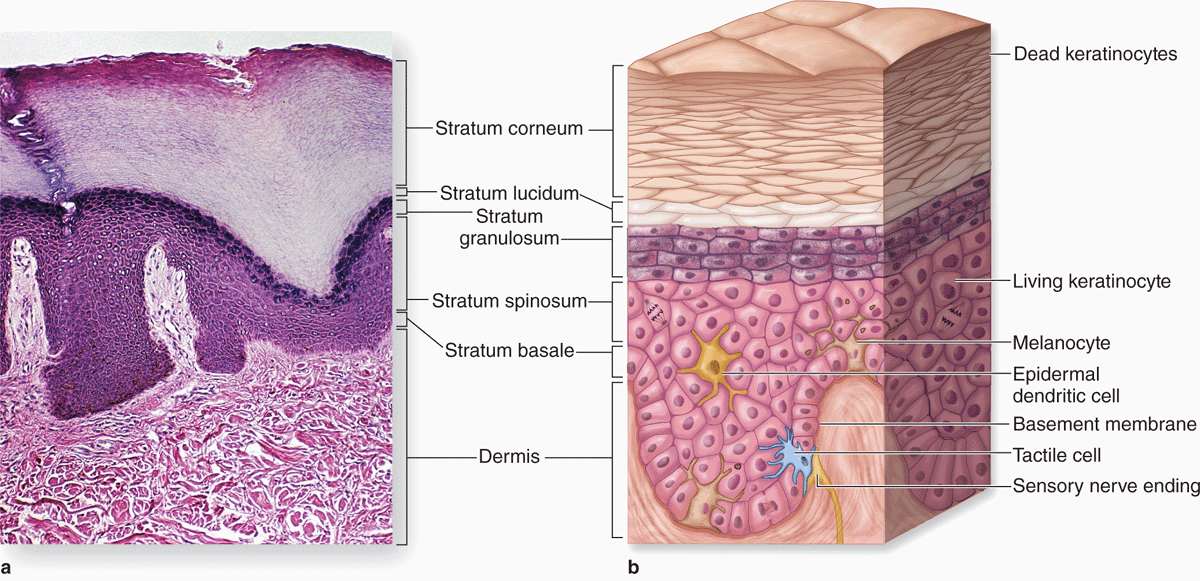
FIGURE 18–2 Layers (strata) of epidermis in thick skin.
The epidermis forms the major distinction between thick skin (Figure 18–2a), found on the palms and soles, and thin skin (Figure 18–3) found elsewhere on the body. The designations “thick” and “thin” refer to the thickness of the epidermal layer, which alone varies from 75 to 150 μm for thin skin and from 400 to 1400 μm (1.4 mm) for thick skin. Total skin thickness (epidermis plus dermis) also varies according to the site. For example, full skin on the back is about 4 mm thick, whereas that of the scalp is about 1.5 mm thick. Like all epithelia, the stratified squamous epidermis lacks microvasculature, its cells receiving nutrients and O2 by diffusion from the dermis.
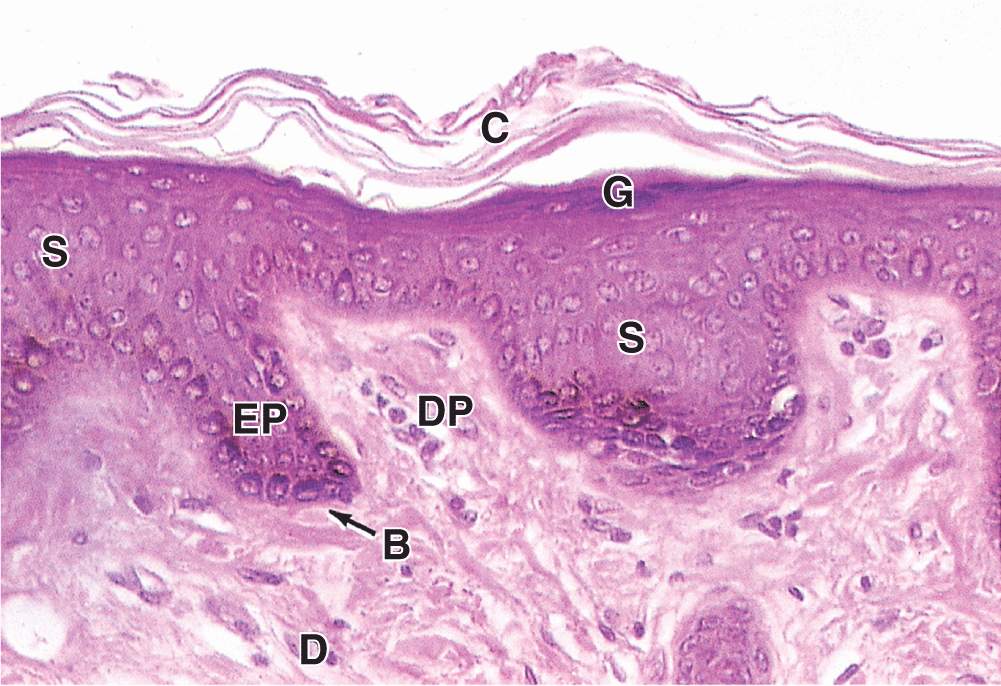
FIGURE 18–3 Layers of epidermis in thin skin.
From the dermis, the epidermis consists of four layers of keratinocytes (or five layers in thick skin, Figure 18–2):
 The basal layer (stratum basale) is a single layer of basophilic cuboidal or columnar cells on the basement membrane at the dermal-epidermal junction (Figures 18–2 and 18–3). Hemidesmosomes in the basal cell membranes join these cells to the basal lamina, and desmosomes bind the cells of this layer together in their lateral and upper surfaces. The stratum basale is characterized by intense mitotic activity and contains, along with the deepest part of the next layer, progenitor cells for all the epidermal layers. In addition to the basal stem cells for keratinocytes found here, a niche for such cells also occurs in the hair follicle sheaths that are continuous with the epidermis. The human epidermis is renewed about every 15 to 30 days, depending on age, the region of the body, and other factors. An important feature of all keratinocytes in the stratum basale is the cytoskeletal keratins, intermediate filaments about 10 nm in diameter. During differentiation, the cells move upward and the amount and types of keratin filaments increase until they represent half the total protein in the superficial keratinocytes.
The basal layer (stratum basale) is a single layer of basophilic cuboidal or columnar cells on the basement membrane at the dermal-epidermal junction (Figures 18–2 and 18–3). Hemidesmosomes in the basal cell membranes join these cells to the basal lamina, and desmosomes bind the cells of this layer together in their lateral and upper surfaces. The stratum basale is characterized by intense mitotic activity and contains, along with the deepest part of the next layer, progenitor cells for all the epidermal layers. In addition to the basal stem cells for keratinocytes found here, a niche for such cells also occurs in the hair follicle sheaths that are continuous with the epidermis. The human epidermis is renewed about every 15 to 30 days, depending on age, the region of the body, and other factors. An important feature of all keratinocytes in the stratum basale is the cytoskeletal keratins, intermediate filaments about 10 nm in diameter. During differentiation, the cells move upward and the amount and types of keratin filaments increase until they represent half the total protein in the superficial keratinocytes.
 MEDICAL APPLICATION
MEDICAL APPLICATION
Friction blisters are lymph-filled spaces created between the epidermis and dermis of thick skin by excessive rubbing, as with ill-fitting shoes or hard use of the hands. If continued, such activity produces protective thickening and hardening of the outer cornified epidermal layers, seen as corns and calluses.
 The spinous layer (stratum spinosum) is normally the thickest layer, especially in the epidermal ridges (Figures 18–2 and 18–3), and consists of generally polyhedral cells having central nuclei with nucleoli and cytoplasm actively synthesizing keratins. Just above the basal layer, some cells may still divide and this combined zone is sometimes called the stratum germinativum. The keratin filaments assemble here into microscopically visible bundles called tonofibrils that converge and terminate at the numerous desmosomes holding the cell layers together. The cells extend slightly around the tonofibrils on both sides of each desmosome (and the extensions elongate if the cells shrink slightly during histologic processing), leading to the appearance of many short “spines” or prickles at the cell surfaces (Figure 18–4). The epidermis of thick skin subject to continuous friction and pressure (such as the foot soles) has a thicker stratum spinosum with more abundant tonofibrils and desmosomes.
The spinous layer (stratum spinosum) is normally the thickest layer, especially in the epidermal ridges (Figures 18–2 and 18–3), and consists of generally polyhedral cells having central nuclei with nucleoli and cytoplasm actively synthesizing keratins. Just above the basal layer, some cells may still divide and this combined zone is sometimes called the stratum germinativum. The keratin filaments assemble here into microscopically visible bundles called tonofibrils that converge and terminate at the numerous desmosomes holding the cell layers together. The cells extend slightly around the tonofibrils on both sides of each desmosome (and the extensions elongate if the cells shrink slightly during histologic processing), leading to the appearance of many short “spines” or prickles at the cell surfaces (Figure 18–4). The epidermis of thick skin subject to continuous friction and pressure (such as the foot soles) has a thicker stratum spinosum with more abundant tonofibrils and desmosomes.
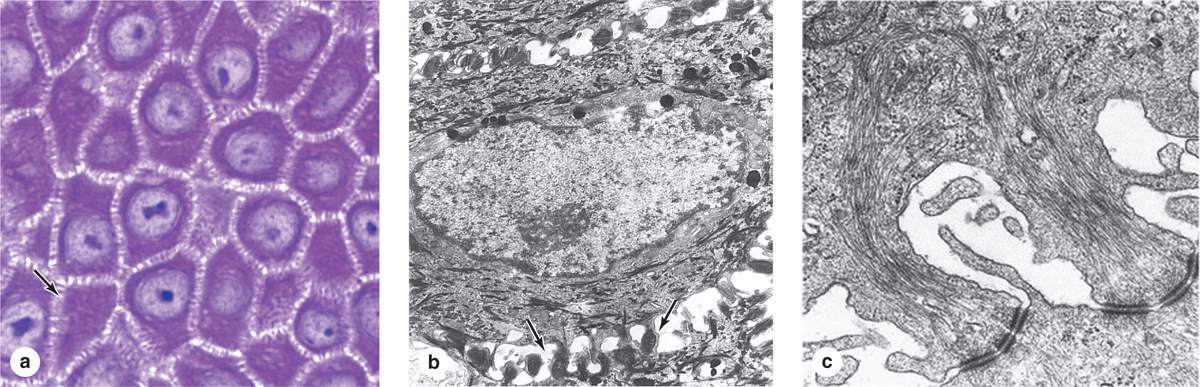
FIGURE 18–4 Keratinocytes of the stratum spinosum,
 MEDICAL APPLICATION
MEDICAL APPLICATION
In adults, one-third of all cancers originate in the skin. Most of these derive from cells of the basal or spinous layers, producing, respectively, basal cell carcinomas and squamous cell carcinomas. Fortunately, both types of tumors can be diagnosed and excised early and consequently are rarely lethal. Skin cancer shows an increased incidence in fair-skinned individuals residing in regions with high amounts of solar radiation.
 The granular layer (stratum granulosum) consists of three to five layers of flattened cells, now undergoing the terminal differentiation process of keratinization. Their cytoplasm is filled with intensely basophilic masses (Figures 18–2 and 18–3; Figure 18–5) called keratohyaline granules. These are dense, non-membrane-bound masses of filaggrin and other proteins associated with the keratins of tonofibrils, linking them further into large cytoplasmic structures. Characteristic ultrastructural features in cells of the granular layer are the membranous, Golgi-derived lamellar granules, small ovoid (100 by 300 nm) structures with many lamellae containing various lipids. Among the last activities of the keratinocytes, the lamellar granules undergo exocytosis, producing a lipid-rich, impermeable layer around the cells. This material forms a major part of the skin’s barrier against water loss. Formation of this barrier, which appeared first in ancestral reptiles, was a key evolutionary process that permitted animals to develop on land. Together, keratinization and production of the lipid-rich layer also have a crucial sealing effect in skin, forming the barrier to penetration by most foreign materials.
The granular layer (stratum granulosum) consists of three to five layers of flattened cells, now undergoing the terminal differentiation process of keratinization. Their cytoplasm is filled with intensely basophilic masses (Figures 18–2 and 18–3; Figure 18–5) called keratohyaline granules. These are dense, non-membrane-bound masses of filaggrin and other proteins associated with the keratins of tonofibrils, linking them further into large cytoplasmic structures. Characteristic ultrastructural features in cells of the granular layer are the membranous, Golgi-derived lamellar granules, small ovoid (100 by 300 nm) structures with many lamellae containing various lipids. Among the last activities of the keratinocytes, the lamellar granules undergo exocytosis, producing a lipid-rich, impermeable layer around the cells. This material forms a major part of the skin’s barrier against water loss. Formation of this barrier, which appeared first in ancestral reptiles, was a key evolutionary process that permitted animals to develop on land. Together, keratinization and production of the lipid-rich layer also have a crucial sealing effect in skin, forming the barrier to penetration by most foreign materials.
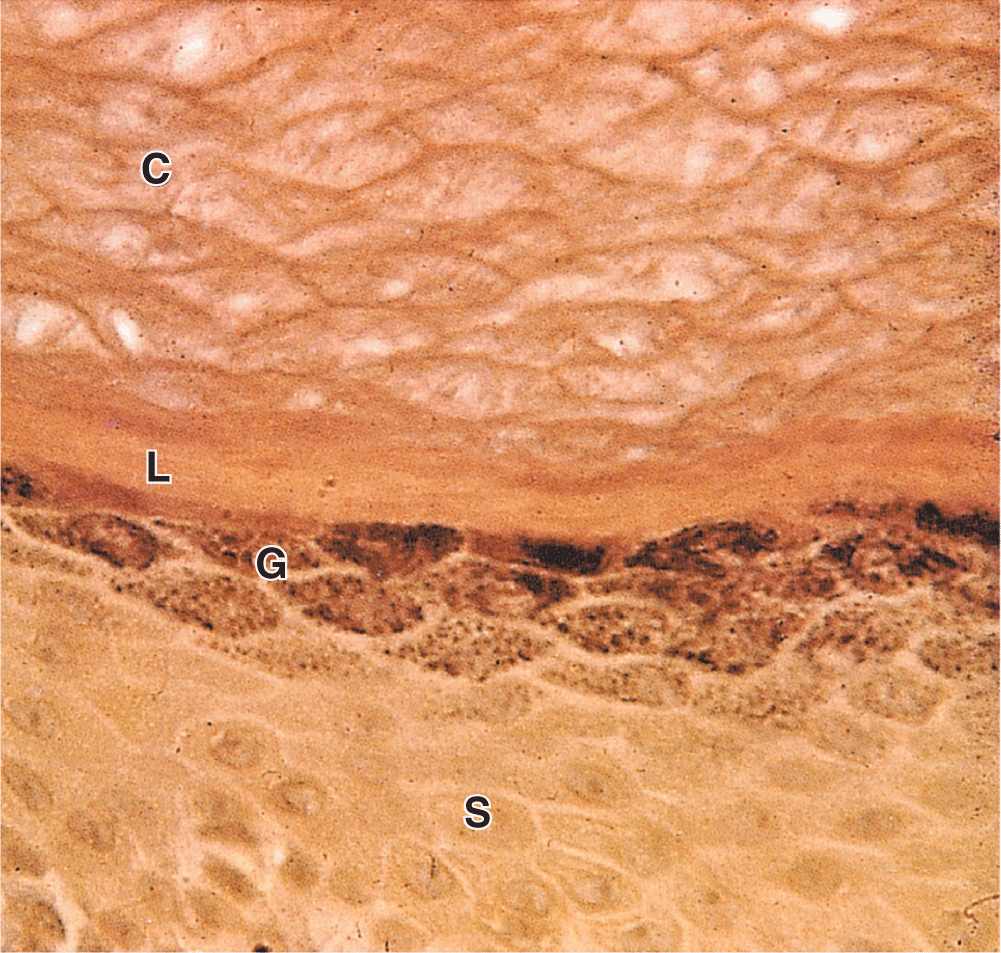
FIGURE 18–5 Stratum granulosum and stratum lucidum: thick skin.
 The stratum lucidum, found only in thick skin, consists of a thin, translucent layer of flattened eosinophilic keratinocytes held together by desmosomes (Figures 18–1 and 18–5). Nuclei and organelles have been lost, and the cytoplasm consists almost exclusively of packed keratin filaments embedded in an electron-dense matrix.
The stratum lucidum, found only in thick skin, consists of a thin, translucent layer of flattened eosinophilic keratinocytes held together by desmosomes (Figures 18–1 and 18–5). Nuclei and organelles have been lost, and the cytoplasm consists almost exclusively of packed keratin filaments embedded in an electron-dense matrix.
 The stratum corneum (Figures 18–2 and 18–3) consists of 15 to 20 layers of squamous, keratinized cells filled with birefringent filamentous keratins. Keratin filaments contain at least six different polypeptides with molecular masses ranging from 40 to 70 kDa, synthesized during cell differentiation in the immature layers. As they form, keratin tonofibrils become heavily massed with filaggrin and other proteins in keratohyaline granules. By the end of keratinization, the cells contain only amorphous, fibrillar proteins with plasma membranes surrounded by the lipid-rich layer. These fully keratinized or cornified cells called squames are continuously shed at the epidermal surface as the desmosomes and lipid-rich cell envelopes break down.
The stratum corneum (Figures 18–2 and 18–3) consists of 15 to 20 layers of squamous, keratinized cells filled with birefringent filamentous keratins. Keratin filaments contain at least six different polypeptides with molecular masses ranging from 40 to 70 kDa, synthesized during cell differentiation in the immature layers. As they form, keratin tonofibrils become heavily massed with filaggrin and other proteins in keratohyaline granules. By the end of keratinization, the cells contain only amorphous, fibrillar proteins with plasma membranes surrounded by the lipid-rich layer. These fully keratinized or cornified cells called squames are continuously shed at the epidermal surface as the desmosomes and lipid-rich cell envelopes break down.
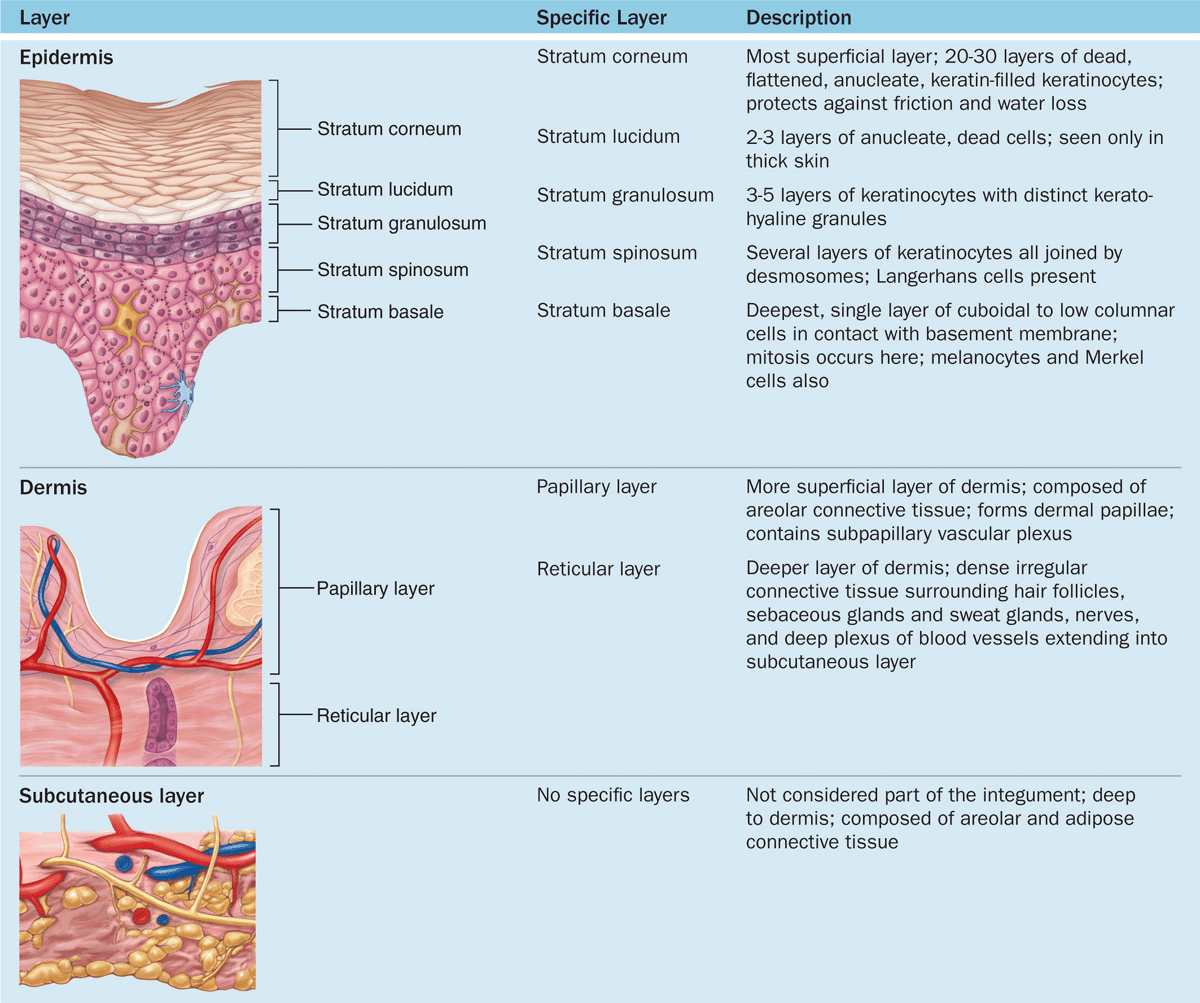
Important features of the epidermal strata are summarized in Table 18–1.
TABLE 18–1 Skin layers and the subcutaneous layer.
 MEDICAL APPLICATION
MEDICAL APPLICATION
In the chronic skin condition called psoriasis, keratocytes are typically produced and differentiate at accelerated rates, causing at least slight thickening of the epidermal layers and increased keratinization and desquamation. Psoriasis is caused by overactive T lymphocytes that trigger an autoimmune reaction in the skin, which can also lead to inflammation with redness, irritation, itching, and scaling, with a defective skin barrier.
Melanocytes
The color of the skin is the result of several factors, the most important of which are the keratinocytes’ content of melanin and carotene and the number of blood vessels in the dermis.
Eumelanins are brown or black pigments produced by the melanocyte (Figures 18–6 and 18–7), a specialized cell of the epidermis found among the cells of the basal layer and in hair follicles. The similar pigment found in red hair is called pheomelanin (Gr. phaios, dusky + melas, black). Melanocytes are neural crest derivatives that migrate into the embryonic epidermis’ stratum basale, where eventually one melanocyte accumulates for every five or six basal keratinocytes (600-1200/mm2 of skin). They have pale-staining, rounded cell bodies attached by hemidesmosomes to the basal lamina, but lacking attachments to the neighboring keratinocytes. Several long irregular cytoplasmic extensions from each melanocyte cell body penetrate the epidermis, running between the cells of the basal and spinous layers and terminating in invaginations of 5 to 10 keratinocytes. Ultrastructurally a melanocyte has numerous small mitochondria, short cisternae of RER, and a well-developed Golgi apparatus (Figure 18–6).
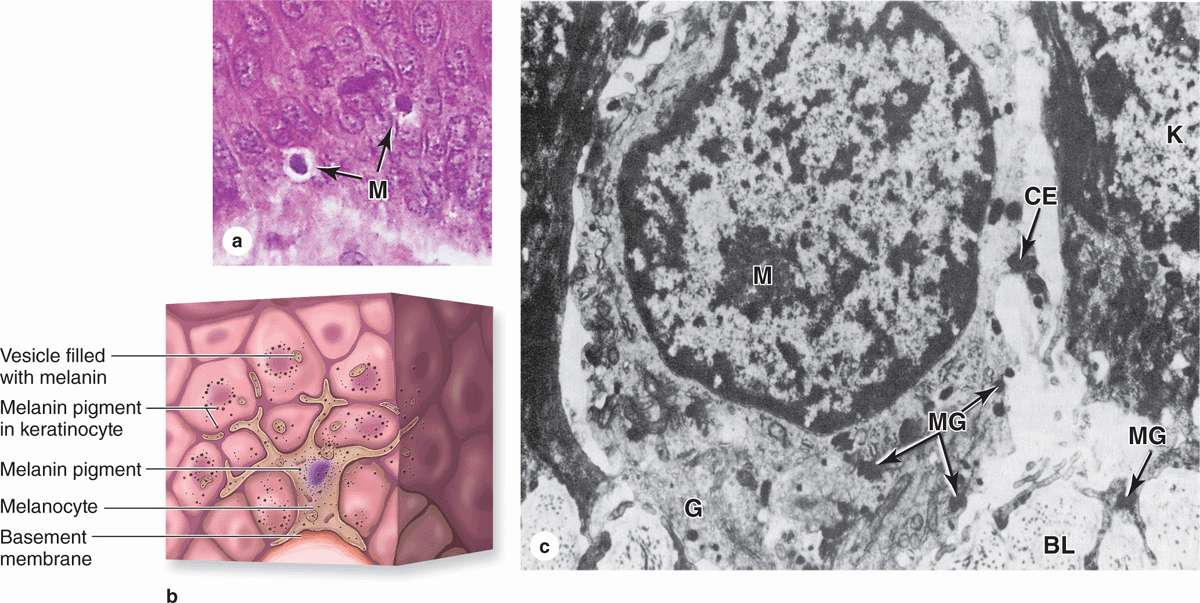
FIGURE 18–6 Melanocytes.
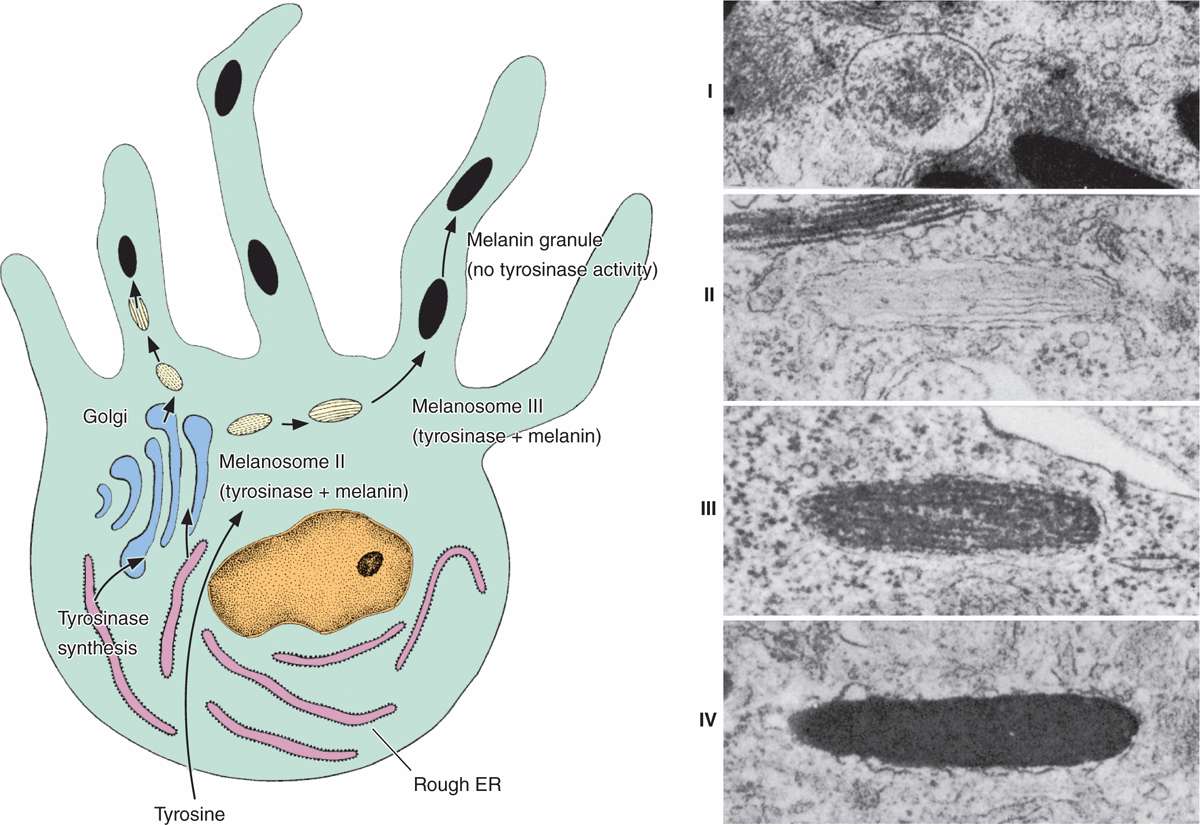
FIGURE 18–7 Melanosome formation.
The first step in melanin synthesis is catalyzed by tyrosinase, a transmembrane enzyme in Golgi-derived vesicles (Figure 18-7a). Tyrosinase activity converts tyrosine into 3,4-dihydroxyphenylalanine (DOPA), which is then further transformed and polymerized into the different forms of melanin. Melanin pigment is linked to a matrix of structural proteins and accumulates in the vesicles until they form mature elliptical granules about 1 μm long called melanosomes (Figure 18–7b).
Melanosomes are then transported via kinesin to the tips of the cytoplasmic extensions. The neighboring keratinocytes phagocytose the tips of these dendrites, take in the melanosomes, and transport them by dynein toward their nuclei. The melanosomes accumulate within keratinocytes as a supranuclear cap that prior to keratinization absorbs and scatters sunlight, protecting DNA of the living cells from the ionizing, mutagenic effects of UV radiation.
Although melanocytes produce melanosomes, the keratinocytes are the melanin depot and contain more of this pigment than the cells that make it. One melanocyte plus the keratinocytes into which it transfers melanosomes make up an epidermal-melanin unit. The density of such units in skin is similar in all individuals. Melanocytes of people with ancestral origins near the equator, where the need for protection against the sun is greatest, produce melanin granules more rapidly and accumulate them more abundantly in keratinocytes.
 MEDICAL APPLICATION
MEDICAL APPLICATION
Albinism is a congenital disorder producing skin hypopigmentation due to a defect in tyrosinase or some other component of the melanin-producing pathway. An acquired condition called vitiligo involves skin depigmentation, often only in affected patches, due to the loss or decreased activity of melanocytes. The causes of melanocyte loss are not clear, but they may include environmental, genetic, or autoimmune conditions.
In regions with much less sunlight such as northern Europe, the small amount of UV radiation penetrating dark skin barely sustains adequate vitamin D3 synthesis. Individuals with ancestry there have one or more genetic polymorphisms that affect steps in melanin formation and cause more lightly pigmented keratinocytes that have increased UV penetration and vitamin D3 synthesis.
Darkening of the skin, or tanning, after exposure to solar radiation at wavelengths of 290 to 320 nm is a two-step process. A physicochemical reaction darkens preexisting melanin. At the same time, paracrine factors secreted by keratinocytes experiencing increased UV radiation accelerate melanin synthesis and its accumulation in the epidermis.



