9 SENSORY ORGANS: VISION AND HEARING
EYE
The eyeball consists of three tunics or layers which, from outside to inside, are (1) the sclera and the cornea, (2) the uvea, and (3) the retina (Figure 9-1).
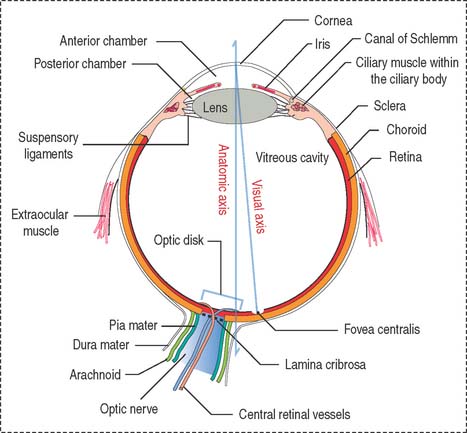
Three distinct and interconnected chambers are found inside the eyeball: the anterior chamber, the posterior chamber, and the vitreous cavity (see Box 9-A). Aqueous humor circulates from the posterior to the anterior chamber. The lens is placed in front of the vitreous cavity, which contains vitreous humor. The bony orbit, the eyelids, the conjunctiva, and the lacrimal apparatus protect the eyeball.
Box 9-A Anatomy of the eye
DEVELOPMENT OF THE EYE
Lateral outpocketings of the right and left sides of the diencephalon give rise to two neuroepithelial optic vesicles, each remaining attached to the brain wall by a hollow optic stalk (Figure 9-2). The surface ectoderm of the head invaginates into the optical vesicle forming a lens vesicle that pinches off. Mesenchyme surrounds both the lens vesicle and the adjacent optic vesicle.
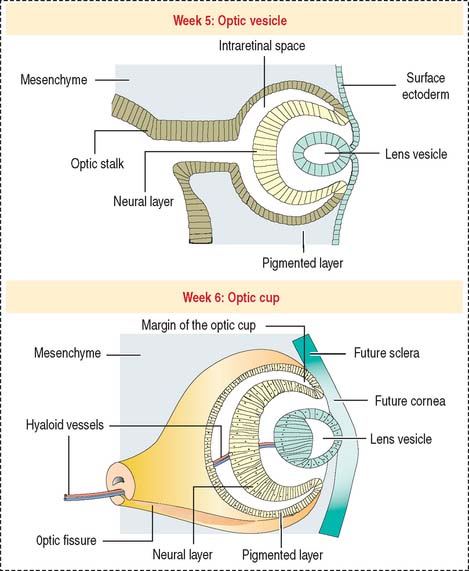
The optic vesicle invaginates and becomes a double-walled optic cup (see Figure 9-2). The optic fissure forms when the outer layer of the optic cup becomes the pigmented epithelium. Cells in the inner layer proliferate and stratify to form the neural retina. The mesenchyme extending into the invagination of the optic cup acquires a gelatinous consistency and becomes the vitreous component of the eye. The lens vesicle is kept in place by the free margins of the optic cup and the surrounding mesenchyme.
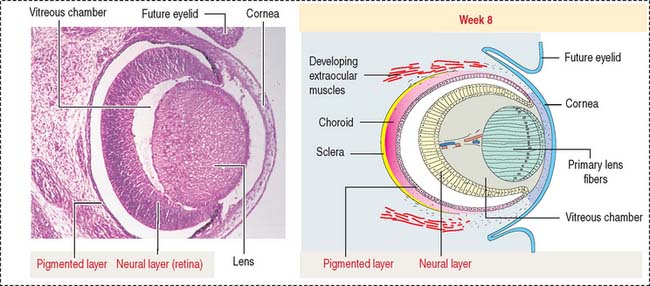
At the outer surface of the optic cup, the mesenchymal shell differentiates into the vascular choroid coat of the eye and the fibrous components of the sclera and cornea (Figure 9-3; see Box 9-B). Posterior to the lens, the vascular choroid coat forms the ciliary body, ciliary muscle, and ciliary processes. Anterior to the lens, the choroid coat forms the stroma of the iris. The ciliary processes secrete the aqueous humor that accumulates first in the posterior chamber (between the iris and lens) and then passes into the anterior chamber (between the lens and cornea) across the pupil. The aqueous humor leaves the anterior chamber by entering into the canal of Schlemm, a small vein (sinus venosus of the sclera) encircling the eye at the anterior edge of the choroid coat.
Box 9-B Development of the cornea
Around the rim of the optic cup, the inner and outer layers form the posterior epithelium of the ciliary body and iris. The sphincter and dilator pupillae muscles develop from the posterior epithelium.
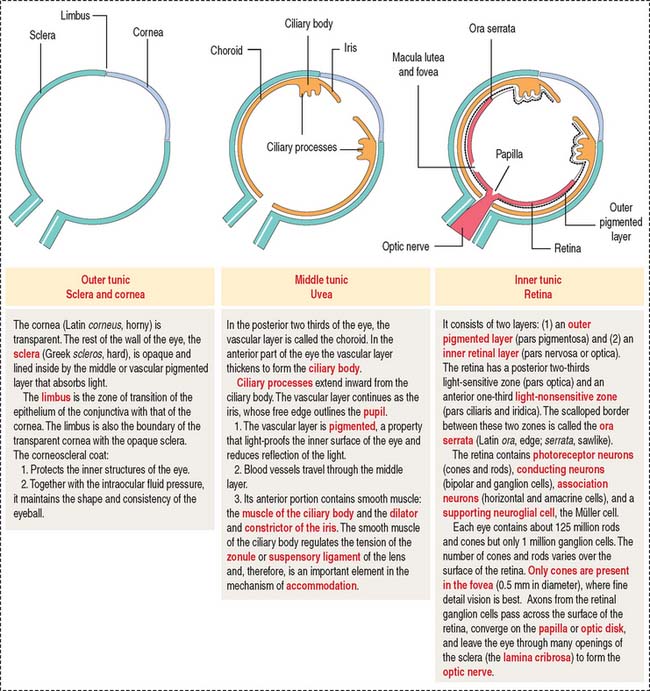
OUTER TUNIC: SCLERA AND CORNEA
The sclera (Figure 9-4) is a 1.0- to 0.4-mm-thick layer of collagen and elastic fibers produced by fibroblasts. The inner side of the sclera faces the choroid, from which it is separated by a layer of loose connective tissue and an elastic tissue network known as the suprachoroid lamina. Tendons of the six extrinsic muscles of the eye are attached to the outer surface of the sclera.
Cornea
The cornea is composed of five layers (Figure 9-5):
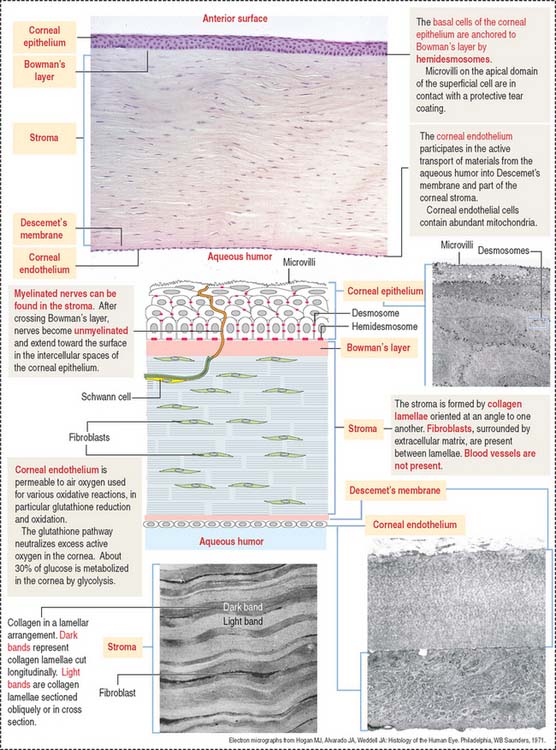
Bowman’s layer is 6 to 9 μm thick, consists of type I collagen fibrils, and lacks elastic fibers. This layer is transparent and does not have regenerative capacity. Bowman’s layer is the anteriormost part of the corneal stroma, although differently organized. For this reason, it is designated “layer” instead of “membrane.” Bowman’s layer represents a protective barrier to trauma and bacterial invasion.
Nerves in transit to the corneal epithelium are found in the corneal stroma.
The corneal endothelium lines the posterior surface of Descemet’s membrane and faces the anterior chamber of the eye. It consists of a single layer of squamous epithelial cells, with impermeable intercellular spaces preventing influx of aqueous humor into the corneal stroma. The structural and functional integrity of the corneal endothelium is vital to the maintenance of corneal transparency (see Box 9-C).
Box 9-C Corneal transplantation
MIDDLE TUNIC: UVEA
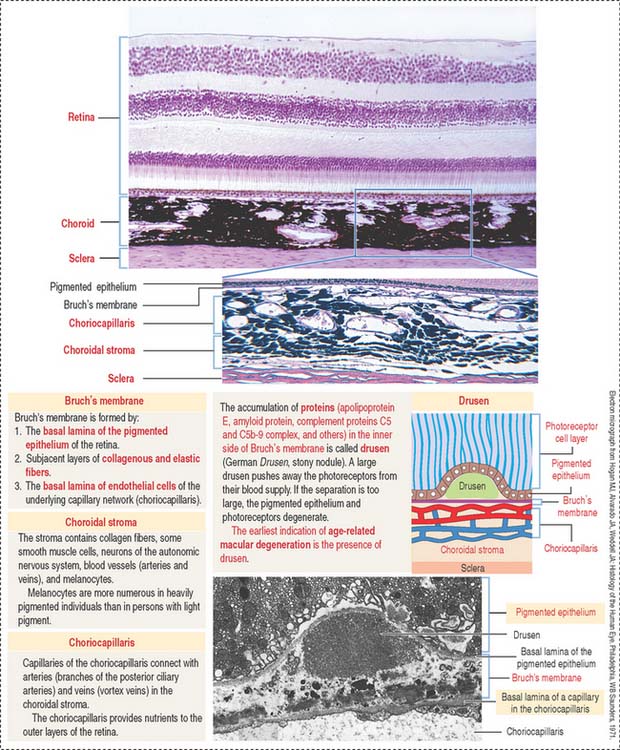
The uvea forms the pigmented vascularized tunic of the eye and is divided into three regions: (1) the choroid, (2) the ciliary body, and (3) the iris (see Figure 9-7) (see Box 9-D).

Box 9-D Uveal tract
The choroid consists of three layers (Figure 9-6):
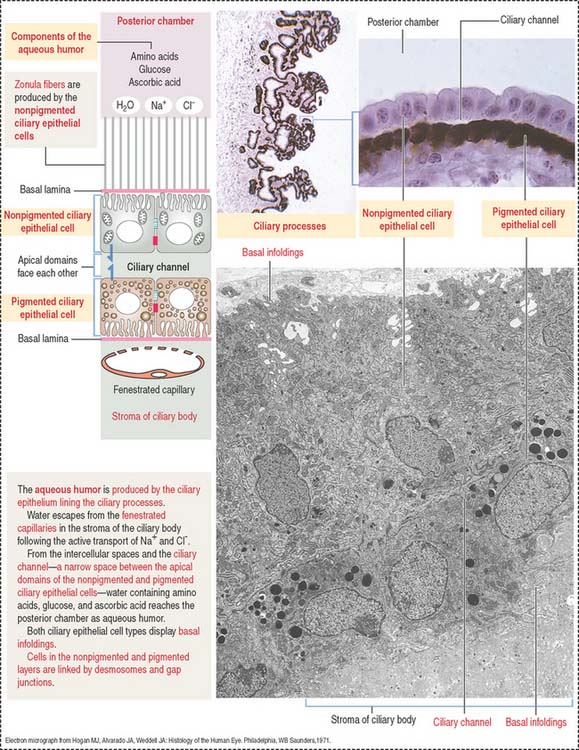
The ciliary body (Figure 9-7) is anterior to the ora serrata and represents the ventral projection of both the choroid and the retina. It is made up of two components: (1) the uveal portion and (2) the neuroepithelial portion.
The uveal portion of the ciliary body includes:
The neuroepithelial portion contributes the two layers of the ciliary epithelium:
Particular features of these two pigmented and nonpigmented epithelial cell layers are:
The anterior (outer) uveal face is of mesenchymal origin and has an irregular surface. It is formed by fibroblasts and pigmented melanocytes embedded in an extracellular matrix. The number of pigmented melanocytes determines the color of the iris. In albinos, the iris appears pink due to the abundant blood vessels. Blood vessels of the iris have a radial distribution and can adjust to changes in length in parallel to variations in the diameter of the pupil.
The posterior (inner) neuroepithelial surface consists of two layers of pigmented epithelium. The outer layer, a continuation of the pigmented layer of the ciliary epithelium, consists of myoepithelial cells that become the dilator pupillae muscle. The smooth muscle of the sphincter pupillae is located in the iris stroma around the pupil.
Three chambers of the eye
The eye contains three chambers (see Figure 9-1): (1) the anterior chamber, (2) the posterior chamber, and (3) the vitreous cavity. The vitreous is the largest component of the eye. The longest part of the optical path from the cornea to the retina is through the vitreous.
The anterior chamber occupies the space between the corneal endothelium (anterior boundary) and the anterior surface of the iris, the pupillary portion of the lens, and the base of the ciliary body (posterior boundary). The circumferential angle of the anterior chamber is occupied by the trabecular meshwork, a drainage site for the aqueous humor into the canal of Schlemm (Figures 9-9 and 9-10).
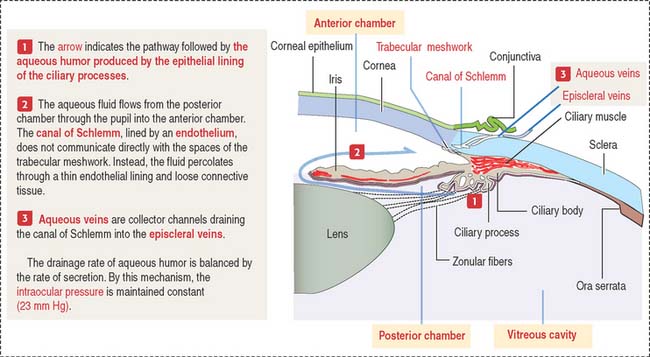
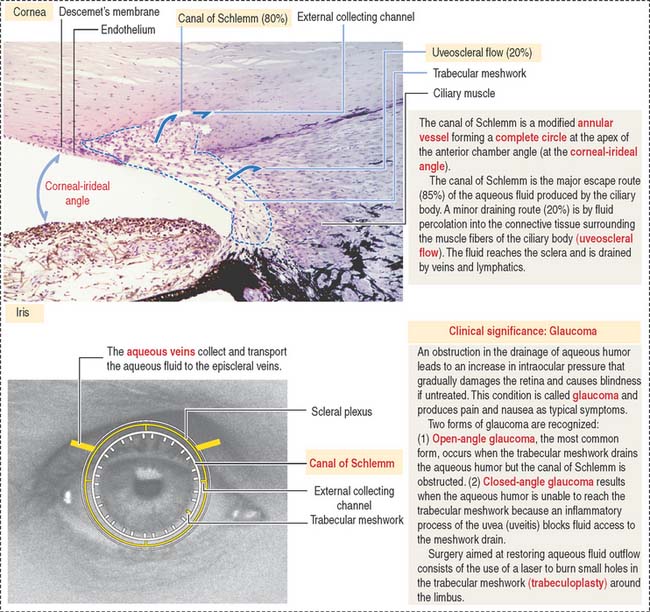
The posterior chamber (see Figure 9-9) is limited anteriorly by the posterior surface of the iris and posteriorly by the lens and the zonular fibers (suspensory ligaments of the lens). The circumferential angle is occupied by the ciliary processes, the site of aqueous humor production.
LENS
The cornea, the three chambers of the eye, and the lens are three transparent structures through which light must pass to reach the retina. Note that the refractive surface of the cornea is an interface between air and tissue and that the lens is in a fluid environment whose refractive index is higher than that of air.
The lens is a transparent, biconvex, elastic, and avascular structure (Figure 9-11). Zonular fibers, consisting of elastin fibrils and a polysaccharide matrix, extend from the ciliary epithelium and insert at the equatorial portion of the capsule. They maintain the lens in place and, during accommodation, change the shape and optical power of the lens in response to forces exerted by the ciliary muscle. The zonular fibers support the lens “as guy wires support a tent.”
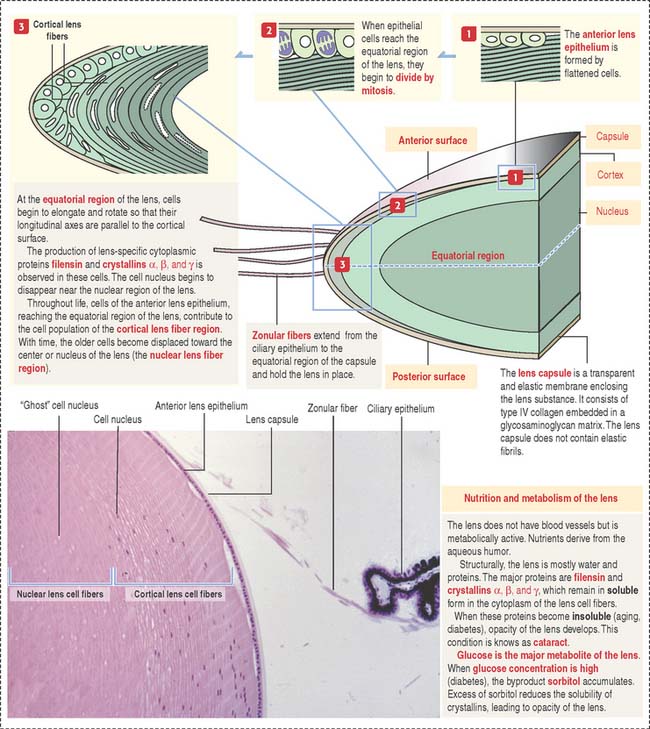
The lens capsule is a thick flexible acellular and transparent basement membrane—like structure containing type IV collagen fibrils and a glycosaminoglycan matrix. Beneath the anterior portion of the capsule is a single layer of cuboidal epithelial cells that extend posteriorly up to the equatorial region. In the cortical region of the lens, elongated and concentrically arranged cells (called cortical lens fibers) arise from the anterior epithelium at the equator region. Cortical lens fibers contain a nucleus and organelles. The nucleus and organelles eventually disappear when the cortical lens fibers approach the center of the lens—the nuclear lens fiber region.
Lens cell differentiation consists of the appearance of unique cytoskeletal proteins: (1) filensin, an intermediate filament that contains attachment sites for crystallins and (2) lens-specific proteins called crystallins (α, β, and γ). Filensin and crystallins maintain the conformation and transparency of the lens fiber cell.
Accommodation
The sharpness of distant and close images focused on the retina depends on the shape of the lens (Figure 9-12). Accommodation defines the process by which the lens becomes rounder to focus the image of a nearby object on the retina and flattens when the image of a distant object is focused on the retina.
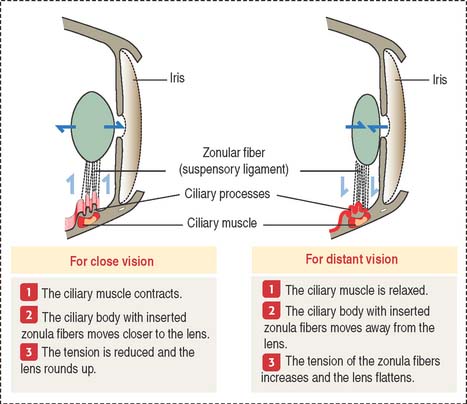
Three components contribute to the accommodation process: (1) the ciliary muscle, (2) the ciliary body, and (3) the suspensory ligaments, inserted at the equatorial region of the lens capsule.
INNER LAYER: RETINA
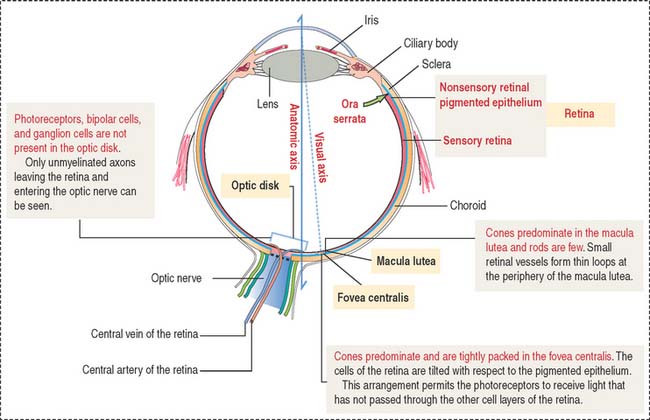
The retina consists of two regions (Figure 9-13): (1) the outer nonsensory retinal pigmented epithelium, and (2) the inner sensory retina (see Box 9-E).
Box 9-E Highlights of the retina
The apical domain of the cuboidal nonsensory pigmented epithelium is sealed by tight junctions to form the external retinal barrier (Figure 9-14). Granules of melanin are present in the apical cytoplasm and apical cell processes. Melanin granules absorb excess light reaching the photoreceptors.
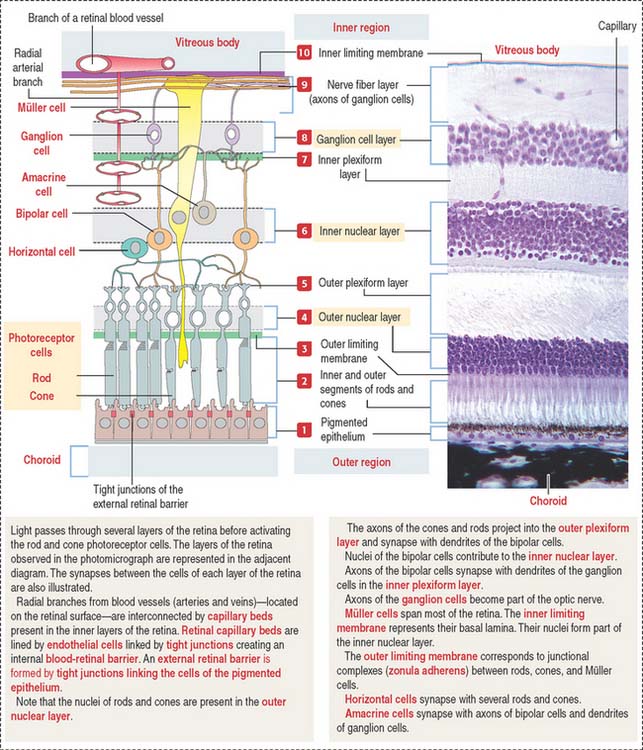
The apical surface contains microvilli that surround the outer segments of the photoreceptors (cones and rods). At this location, the sensory retina and the pigmented epithelium are attached to each other through an amorphous extracellular material, the interphotoreceptor matrix (Figure 9-15).
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


