Recognizing and Classifying Cells
Light microscopic examination of stained cells in smears is the method of choice of diagnostic cytology. It allows classification of most normal cells as to type and tissue of origin. It also allows the recognition of cell changes caused by disease processes, discussed in general terms in Chapters 6 and 7 and, more specifically, in subsequent chapters.
GENERAL GUIDELINES
The study of cells in smears should take place at several levels:
A rapid review of the smear with a 10× objective provides information on the makeup of the sample and its cell content. This preliminary review will tell the observer whether the smear is appropriately fixed and stained and will provide initial information on its composition. Smears containing only blood or no cells at all are usually considered inadequate, with some very rare exceptions.
If the smear contains cells other then blood cells, it should be examined with care. A careful review of the material or screening of smears with a 10× objective is usually required to identify abnormal cells that may be few in number. Screening is mandatory in cancer detection samples from “well” patients. A microscope stage should be utilized. The methods of screening are described in Chapter 44.
The screening of the smear should lead to the preliminary assessment of the sample and answer the following questions: (1) Does the cell population correspond to the organ of origin? (2) If the answer is positive, the next question pertains to the status of the cell population: (a) is it normal? (b) does it show nonspecific abnormalities of little consequence to the patient? or (c) Does it show abnormalities pertaining to a recognizable disease state that can be identified?
To answer these questions, fundamental principles of cell classification must be presented.
CELL CLASSIFICATION
An Overview of the Problem
In general, the derivation, type of cells, and sometimes their function, are reflected in the cytoplasm, whereas the nucleus offers information on the status of the DNA, which is of particular value in the diagnosis of cancer. Some cells that lack distinct cytoplasmic or nuclear features may be very difficult to classify. Nuclear and nucleolar changes in cancer are described in detail in Chapter 7.
Knowledge of the rudiments of histology is necessary for cell classification. For all practical purposes, the cells encountered in cytologic samples are of epithelial and nonepithelial origin. The most common cell types will be discussed here. Other cell types will be described as needed in appropriate chapters.
With the development of monoclonal or polyclonal antibodies to specific cell components, still further insights into cell derivation and function can be achieved by immunocytochemistry. An immunochemical analysis of the components of the cell skeleton, such as the intermediate filaments, of cell products, such as various hormones, and of immunologic features vested in the cell membrane, allows additional analysis and classification of cells (see Chap. 45).
An additional point must be made in reference to the comparison of tissue sections and cells of the same origin.
In tissue sections, the cells are often cut “on edge” and are seen in profiles. In cytologic preparations, the cells are whole and are generally flattened on a glass slide, usually affording a much better analysis of the cell components. A schematic comparison of histology and cytology is shown in Figure 5-1. A description of the principal tissue and cell types observed in diagnostic cytology is provided below.
In tissue sections, the cells are often cut “on edge” and are seen in profiles. In cytologic preparations, the cells are whole and are generally flattened on a glass slide, usually affording a much better analysis of the cell components. A schematic comparison of histology and cytology is shown in Figure 5-1. A description of the principal tissue and cell types observed in diagnostic cytology is provided below.
Epithelial Cells
An epithelium (plural: epithelia) is a tissue lining the surfaces of organs or forming glands and gland-like structures. Similar epithelia may occur in various organs and organ systems. There are four principal groups of epithelia: (1) squamous epithelia, synonymous with protective function; (2) glandular epithelia with secretory functions; (3) ciliated epithelia; and (4) the mesothelia.
Squamous Epithelium
Histology
The squamous epithelium is a multilayered epithelium that lines the surfaces of organs that are in direct contact with the external environment. Two subtypes of this epithelium can be recognized: the keratinizing type, occurring in the skin and the outer surface of the vulva and the non-keratinizing type, occurring in the buccal cavity, cornea, pharynx, esophagus, vagina and the inner surface of the vulva, and the vaginal portio of the cervix. The differences between the two subtypes of squamous epithelium reside in their mechanisms of maturation and formation of the superficial layers, discussed below.
Squamous epithelium is organized in multiple layers. Starting at the bottom of the epithelium, resting on the lamina propria, to the top of the epithelium, facing the surface, four principal layers can be distinguished, although the separation of the layers is arbitrary. The bottom, basal layer, is composed of small cells. Immediately above are the parabasal layers, composed of two or three layers of somewhat larger cells, which blend with the next intermediate layers, composed of several layers of larger cells. The fourth superficial layers of the squamous epithelium are composed of a variable number of layers of the largest cells.
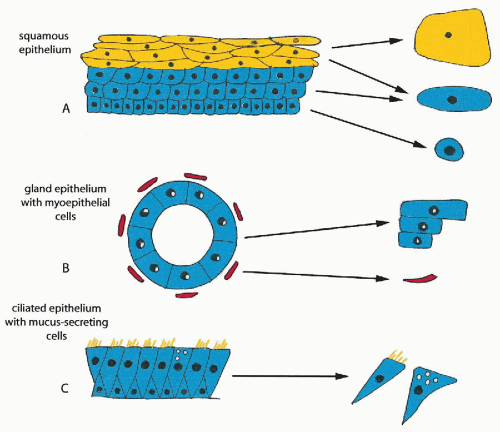 Figure 5-1 Comparison of histology and cytology of three common types of epithelia. A. Squamous epithelium. The cells are provided with a rigid skeleton of intermediate filaments; hence, they are resistant to injury. The cells vary in size and configuration, depending on the layer of origin. Cells from the superficial layer are large, with abundant cytoplasm and small nuclei. Cells from the intermediate and parabasal layers are smaller and have an open, spherical nuclei (vesicular nuclei). Cells from the basal layer are still smaller, but the nuclear structure is identical with that of parabasal cells. B. Glandular epithelium. These epithelia are usually quite fragile and are often injured, hence, poorly preserved. The cells may vary in size from cuboidal to columnar. Small contractile myoepithelial cells often accompany glandular cells. C. Ciliated epithelium with mucus-secreting cells. The ciliated cells are readily recognized because of the flat, cilia-bearing surface and a thing, taillike opposite end. The mucus-producing cells (goblet cells) are of a similar configuration but have no cilia, and their cytoplasm is distended with mucus-containing vacuoles. |
The epidermis of the skin is the prototype of squamous epithelium (Fig. 5-2). The features conferring special strength on this epithelium are keratin filaments of high relative molecular mass, and numerous desmosomes, cell junctions that are very difficult to disrupt (Fig. 5-3; see also Fig. 2-13).
The growth of the squamous epithelium is in the direction of the surface, that is, the cells move from the basal layer, to parabasal layers, to intermediate layers, to superficial layers. The most superficial cells are cast off. Under conditions of health, the small cells of the basal layer are the only cells in this type of epithelium that are capable of mitosis. It should be noted that the cells of the basal layer have several different functions: some anchor the epithelium to the basement lamina, some provide new basal cells to ensure the survival of the epithelium, and some produce cells that are destined to mature and thus form the bulk of the epithelium. There are no morphologic differences among the basal cells with different functions.
As the cells transit from the basal to the more superficial layers, they are programmed to gradually increase the size of their cytoplasm. The increase in the size of the cytoplasm is accompanied by an increase in the intermediate keratin filaments of high relative molecular weight (see Fig. 2-27). As the cells progress through the stages of maturation, they are bound to each other by desmosomes, until they reach the superficial layer, where the desmosomes disintegrate to allow shedding of the most superficial cells. The process of cytoplasmic maturation is accompanied by nuclear changes. The nuclei of the basal, parabasal, and intermediate layers of squamous cells appears as spherical, open (vesicular) structures, measuring approximately 8 μm in diameter. As the cells transit from the intermediate to superficial layers, their nuclei shrink and become condensed (nuclear pyknosis).
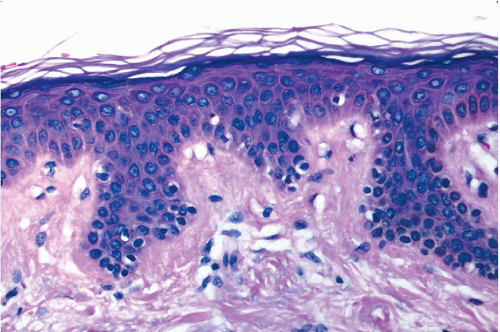 Figure 5-2 Histologic section of normal human skin as an example of squamous epithelium with protective function. Note the small cuboidal cells of the basal layer adjacent to connective tissue of the dermis (bottom). The surface is formed by several “basketweave” layers of anucleated squames. The bulk of the epithelium is composed of intermediate cells. Scattered cells with clear cytoplasm are the Langerhans’ cells, representing the immune system. |
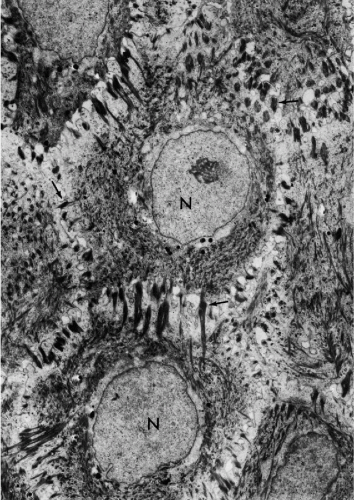 Figure 5-3 Electron micrograph of middle layer of human epidermis. The nuclei (N) are surrounded by a perinuclear clear zone free of filaments. The remaining cytoplasm shows an abundance of intermediate filaments forming aggregates (bundles) seen in longitudinal, oblique, or transverse section. Many of the filament bundles terminate on the numerous desmosomes, some identified by arrows. The integrity of the desmosomes accounts for the cohesion of this type of epithelium. (× 18,000.) (Courtesy of the late Dr. Philip Prose, New York University, New York.) |
The differences between the two subtypes of the squamous epithelium are evident in the superficial layers: in the nonkeratinizing squamous epithelium, the superficial cells are cast off, while still retaining their nuclei (see Chaps. 8 and 19). In the keratinizing squamous epithelium, such as the epidermis of the skin, the superficial cells continue to accumulate keratin filaments, which obliterate the nucleus until the cell becomes an anucleated, keratinfilled shell (anucleated squames). The anucleated squames of the epidermis form a superficial horny layer, which provides the best protection against injury (see Fig. 5-2).
Under abnormal circumstances, formation of a horny layer may also occur in nonkeratinizing squamous epithelia, resulting in white patches visible with the naked eye, and, therefore, known as leukoplakia (from Latin, leukos = white and plax = plaque). This condition may occur in the uterine cervix or the buccal cavity and is described in the appropriate chapters.
Squamous epithelia are also provided with cells with immune function, the Langerhans’ cells, characterized by clear, transparent cytoplasm (see Fig. 5-2). These cells appear to mediate a broad variety of immunologic responses of the squamous epithelia to environmental and internal stimuli (summary in Robert and Kupper, 1999).
Cytology
Cells derived from squamous epithelia are usually quite resilient to manipulation and often retain their shape because of high keratin content. In general, these cells tend to be flat, polygonal, and sharply demarcated, and they vary in size according to the layer of origin. The smallest cells, measuring about 10 μm in diameter, are the basal cells, which are very rarely seen in normal states. Parabasal cells, derived from the parabasal layers, are somewhat larger, measuring from 10 to 15 μm in diameter. Intermediate cells, derived from the intermediate layers, are still larger, measuring from 15 to 40 μm in diameter. The superficial cells are the largest, measuring from 40 to 60 μm in diameter. The cells derived from the basal, parabasal, and intermediate layers show spherical nuclei, resembling open vesicles, with delicate chromatin, hence the term vesicular nuclei, measuring about 8 μm in diameter. The superficial squamous cells derived from non-keratinizing squamous epithelium, show small, condensed, and dark nuclei that are often encircled by a narrow clear cytoplasmic zone of contraction. Such nuclei are referred to as pyknotic nuclei (from Greek, pyknos = dense) (Figs. 5-4 and 5-5). Anucleated squames, derived from keratinizing squamous epithelium, appear as polygonal, transparent structures without visible nuclei. The staining characteristics of the cytoplasm in cytologic preparations presumably depends on the species of keratin filaments. The cytoplasm of the superficial cells is usually eosinophilic. The cytoplasm of cells from the lower cell layers is usually basophilic. These staining properties may be modified by exposure to air-drying, which often results in a tinctorial change from basophilic to eosinophilic.
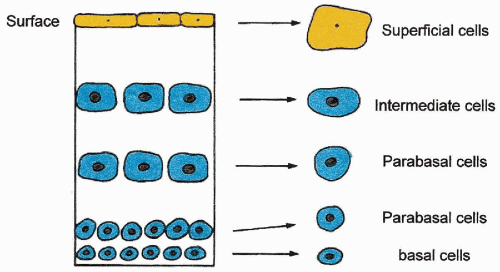 Figure 5-4 Diagrammatic representation of a squamous epithelium (other than epidermis of the skin), comparing the morphologic designation of cell types and their derivation from various epithelial layers. |
Other Protective Epithelia
Variants of squamous epithelium, often highly specialized, may be observed in a variety of organ systems, for example, in the lower urinary tract and the larynx. The special features of these epithelia and the cells derived therefrom are described in the appropriate chapters.
Epithelia With Secretory Function
Histology
These epithelia are found mainly in organs with secretory functions and exchanges with the external environment,
such as food intake, principally in the digestive tract and associated glands. Similarly structured epithelia also occur in other locations, such as the male and female genital tracts. Secretory epithelia that line the surfaces of organs, such as the intestine and the endocervix, form invaginations or crypts, or may be organized in glands connected with the surface by ducts. Single cells of secretory type may also occur as a component of other epithelial types, for example, as goblet cells in the ciliated epithelium of the respiratory tract (see Fig. 5-1).
such as food intake, principally in the digestive tract and associated glands. Similarly structured epithelia also occur in other locations, such as the male and female genital tracts. Secretory epithelia that line the surfaces of organs, such as the intestine and the endocervix, form invaginations or crypts, or may be organized in glands connected with the surface by ducts. Single cells of secretory type may also occur as a component of other epithelial types, for example, as goblet cells in the ciliated epithelium of the respiratory tract (see Fig. 5-1).
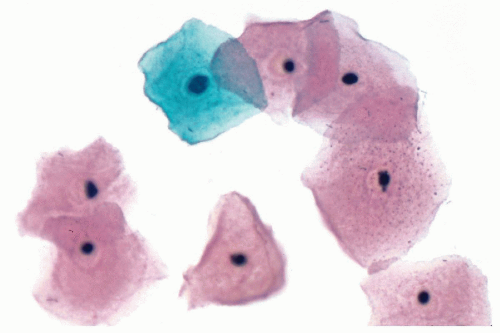 Figure 5-5 Mature squamous cells characterized by production of a large, resilient cytoplasmic surface. The condensed (pyknotic) nucleus is comparatively small. This cell type is eminently suited for the exercise of protective function. (Human buccal epithelium.) |
Secretory epithelia are usually made up of a single layer of cuboidal or columnar cells with a clear or opaque cytoplasm and vesicular nuclei (see Figs. 5-1 and 5-6). The nuclei are often located at the periphery of the cells, away from the lumen of the organ. The cytoplasm contains the products of cell secretion, such as mucus. The replacements for such epithelial cells are provided by small, intercalated basal cells (reserve cells), which, under circumstances not clearly defined, replace obsolete glandular cells. The third component of secretory epithelia observed only in glands and ducts, such as salivary glands and ducts, is a peripheral layer of elongated cells with contractile properties, known as the myoepithelial cells (see Fig. 5-1). The function of the myoepithelial cells is to propel the product of cell secretions into excretory ducts and beyond.
Ultrastructural features of secretory epithelia were discussed in Chapter 2. The cells are provided with a large Golgi apparatus wherein the synthesis of the products of secretion takes place. The superficial cells form tight junctions that protect the internal environment of such epithelia.
Cytology
When well preserved, the secretory cells are cuboidal or columnar in shape, averaging from 10 to 20 μm in length and 10 μm in width. Their cytoplasm is transparent because of accumulation of products of secretion, usually mucus (Fig. 5-7). The products of secretion are packaged in small cytoplasmic vacuoles. It is important to note that secretory cells are often polarized, that is, they display one flat surface facing the lumen of the organ. Through that surface, the cells products are discharged. The nuclei of the secretory cells are open (vesicular), averaging about 8 μm in diameter. The nuclei are either clear (transparent) or show moderate granularity, and are often provided with small nucleoli. The cytoplasm of cells derived from secretory epithelia is fragile and difficult to preserve. Thus, when these cells are removed from their site of origin, they often have poorly demarcated borders and their shape may be distorted. The cytoplasm of most secretory cells accepts pale basophilic stains.
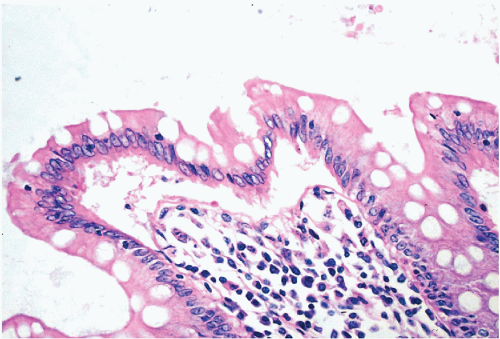 Figure 5-6 Columnar epithelium of normal human colon. Note the opaque columnar cells and very many clear goblet cells. (H & E.) |
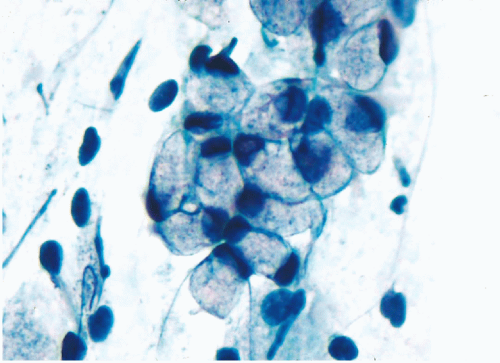 Figure 5-7 Mucus-secreting endocervical cells. The cytoplasm of these cells is filled with mucus, which remains unstained. The nuclei are pushed to the periphery. Compare with electron micrographs of somewhat similar cells (see Figs. 1-15 and 1-19). This is a good example of a glandular cell with the cytoplasmic features geared to excretory function. |
The myoepithelial cells are seen only in aspirated samples and are recognized by their small, comma-shaped, dark nuclei, surrounded by a very narrow rim of cytoplasm (see Chap. 29).
Ciliated Epithelia
Histology
The ciliated epithelia are characterized by columnar, rarely cuboidal cells with one ciliated surface that is facing the lumen of the organ. Such cells occur mainly in the respiratory tract, where they line the bronchi (see Fig. 1-4 and Chap. 19) but may be also found in the endocervix, the fallopian tube, and the endometrium during the secretory phase. As an incidental finding, ciliated cells may be occasionally observed in almost any secretory epithelium. Very often, the ciliated cells are accompanied by secretory cells that produce mucus or related substances, for example, goblet cells in the respiratory tract (see Fig. 5-1). The ciliated epithelia are often stratified,




Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
