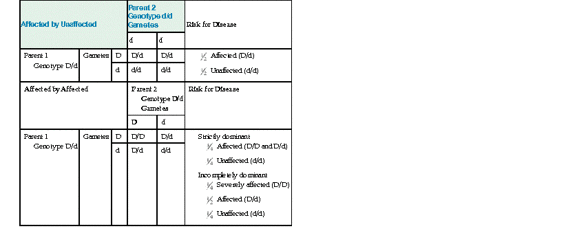
As seen in the table, when both parents of an affected person are carriers, their children’s risk for receiving a recessive allele is 50% from each parent. The chance of inheriting two recessive alleles and therefore being affected is thus 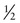 ×
× 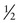 or 1 in 4 with each pregnancy. The 25% chance for two heterozygotes to have a child with an autosomal recessive disorder is independent of how many previous children there are who are either affected or unaffected. The proband may be the only affected family member, but if any others are affected, they are usually in the same sibship and not elsewhere in the kindred (Fig. 7-4).
or 1 in 4 with each pregnancy. The 25% chance for two heterozygotes to have a child with an autosomal recessive disorder is independent of how many previous children there are who are either affected or unaffected. The proband may be the only affected family member, but if any others are affected, they are usually in the same sibship and not elsewhere in the kindred (Fig. 7-4).
Sex-Influenced Autosomal Recessive Disorders
Because males and females both have the same complement of autosomes, autosomal recessive disorders generally show the same frequency and severity in males and females. There are, however, exceptions. Some autosomal recessive diseases demonstrate a sex-influenced phenotype, that is, the disorder is expressed in both sexes but with different frequencies or severity. For example, hereditary hemochromatosis is an autosomal recessive phenotype that is 5 to 10 times more common in males than in females (Case 20). Affected individuals have enhanced absorption of dietary iron that can lead to iron overload and serious damage to the heart, liver, and pancreas. The lower incidence of the clinical disorder in homozygous females is believed to be due to their lower dietary iron intake, lower alcohol usage, and increased iron loss through menstruation.
Gene Frequency and Carrier Frequency
Mutant alleles responsible for a recessive disorder are generally rare, and so most people will not have even one copy of the mutant allele. Because an autosomal recessive disorder must be inherited from both parents, the risk that any carrier will have an affected child depends partly on the chance that his or her mate is also a carrier of a mutant allele for the condition. Thus knowledge of the carrier frequency of a disease is clinically important for genetic counseling.
The most common autosomal recessive disorder in white children is cystic fibrosis (CF) (Case 12), caused by mutations in the CFTR gene (see Chapter 12). Among white populations, approximately 1 child in 2000 has two mutant CFTR alleles and has the disease, from which we can infer that 1 in 23 individuals is a silent carrier who has no disease. (How one calculates heterozygote frequencies in autosomal recessive conditions will be addressed in Chapter 9.) Mutant alleles may be handed down from carrier to carrier for numerous generations without ever appearing in the homozygous state and causing overt disease. The presence of such hidden recessive genes is not revealed unless the carrier happens to mate with someone who also carries a mutant allele at the same locus and the two deleterious alleles are both inherited by a child.
Estimates of the number of deleterious alleles in each of our genomes range from 50 to 200 based on examining an individual’s complete exome or genome sequence for clearly deleterious mutations in the coding regions of the genome (see Chapter 4). This estimate is imprecise, however. It may be an underestimate, because it does not include mutant alleles whose deleterious effect is not obvious from a simple examination of the DNA sequence. It may also, however, be an overestimate, because it includes mutations in many genes that are not known to cause disease.
Consanguinity
Because most mutant alleles are generally uncommon in the population, people with rare autosomal recessive disorders are typically compound heterozygotes rather than true homozygotes. One well-recognized exception to this rule occurs when an affected individual inherits the exact same mutant allele from both parents because the parents are consanguineous (i.e., they are related and carry the identical mutant allele inherited from a common ancestor). Finding consanguinity in the parents of a patient with a genetic disorder is strong evidence (although not proof) for the autosomal recessive inheritance of that condition. For example, the disorder in the pedigree in Figure 7-5 is likely to be an autosomal recessive trait, even though other information in the pedigree may seem insufficient to establish this inheritance pattern.
Consanguinity is more frequently found in the background of patients with very rare conditions than in those with more common recessive conditions. This is because it is less likely that two individuals mating at random in the population will both be carriers of a very rare disorder by chance alone than it is that they would both be carriers because they inherited the same mutant allele from a single common ancestor. For example, in xeroderma pigmentosum (Case 48), a very rare autosomal recessive condition of DNA repair (see Chapter 15), more than 20% of cases occur among the offspring of marriages between first cousins. In contrast, in more common recessive conditions, most cases of the disorder result from matings between unrelated persons, each of whom happens by chance to be a carrier. Thus most affected persons with a relatively common disorder, such as CF, are not the result of consanguinity, because the mutant allele is so common in the general population. How consanguinity is measured for different matings is described in Chapter 9.
The genetic risk to the offspring of marriages between related people is not as great as is sometimes imagined. For marriages between first cousins, the absolute risks of abnormal offspring, including not only known autosomal recessive diseases but also stillbirth, neonatal death, and congenital malformation, is 3% to 5%, approximately double the overall background risk of 2% to 3% for offspring born to any unrelated couple (see Chapter 16). Consanguinity at the level of third cousins or more remote relationships is not considered to be genetically significant, and the increased risk for abnormal offspring is negligible in such cases.
The incidence of first-cousin marriage is low (≈1 to 10 per 1000 marriages) in many populations in Western societies today. However, it remains relatively common in some ethnic groups, for example, in families from rural areas of the Indian subcontinent, in other parts of Asia, and in the Middle East, where between 20% and 60% of all marriages are between cousins.
Characteristics of Autosomal Recessive Inheritance
• An autosomal recessive phenotype, if not isolated, is typically seen only in the sibship of the proband, and not in parents, offspring, or other relatives.
• For most autosomal recessive diseases, males and females are equally likely to be affected.
• Parents of an affected child are asymptomatic carriers of mutant alleles.
• The parents of the affected person may in some cases be consanguineous. This is especially likely if the gene responsible for the condition is rare in the population.
• The recurrence risk for each sib of the proband is 1 in 4 (25%).
Autosomal Dominant Inheritance
More than half of all known mendelian disorders are inherited as autosomal dominant traits. The incidence of some autosomal dominant disorders can be high. For example, adult polycystic kidney disease (Case 37) occurs in 1 in 1000 individuals in the United States. Other autosomal dominant disorders show a high frequency only in certain populations from specific geographical areas: for example, the frequency of familial hypercholesterolemia (Case 16) is 1 in 100 for Afrikaner populations in South Africa and of myotonic dystrophy is 1 in 550 in the Charlevoix and Saguenay–Lac Saint Jean regions of northeastern Quebec. The burden of autosomal dominant disorders is further increased because of their hereditary nature; when they are transmitted through families, they raise medical and even social problems not only for individuals but also for whole kindreds, often through many generations.
The risk and severity of dominantly inherited disease in the offspring depend on whether one or both parents are affected and whether the trait is a pure dominant or is incompletely dominant. There are a number of different ways that one mutant allele can cause a dominantly inherited trait to occur in a heterozygote despite the presence of a normal allele. Disease mechanisms in various dominant conditions are discussed in Chapter 12.
Denoting D as the mutant allele and d as the wild-type allele, matings that produce children with an autosomal dominant disease can be between two heterozygotes (D/d) for the mutation or, more frequently, between a heterozygote for the mutation (D/d) and a homozygote for a normal allele (d/d).
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


