Structure & Function in the Liver
The organs associated with the digestive tract include the major salivary glands, the pancreas, the liver, and the gallbladder. Products of these organs facilitate transport and digestion of food within the gastrointestinal tract. The main functions of the salivary glands are to moisten and lubricate ingested food and the oral mucosa, to initiate the digestion of carbohydrates and lipids with amylase and lipase, and to secrete innate immune components such as lysozyme and lactoferrin.
The pancreas secretes digestive enzymes that act in the small intestine and hormones important for the metabolism of the absorbed nutrients. Bile, whose components are necessary for digestion and absorption of fats, is made in the liver but stored and concentrated in the gallbladder. The liver also plays a major role in carbohydrate and protein metabolism, inactivates many toxic substances and drugs, and synthesizes most plasma proteins and factors necessary for blood coagulation.
SALIVARY GLANDS
Exocrine glands in the mouth produce saliva, which has digestive, lubricating, and protective functions. With a normal pH of 6.5 to 6.9, saliva also has an important buffering function and in some species is also important for evaporative cooling. There are three pairs of large salivary glands: the parotid, submandibular, and sublingual glands (Figure 16–1), in addition to the numerous minor mucosal salivary glands throughout the oral cavity, which secrete about 10% of the total saliva volume.
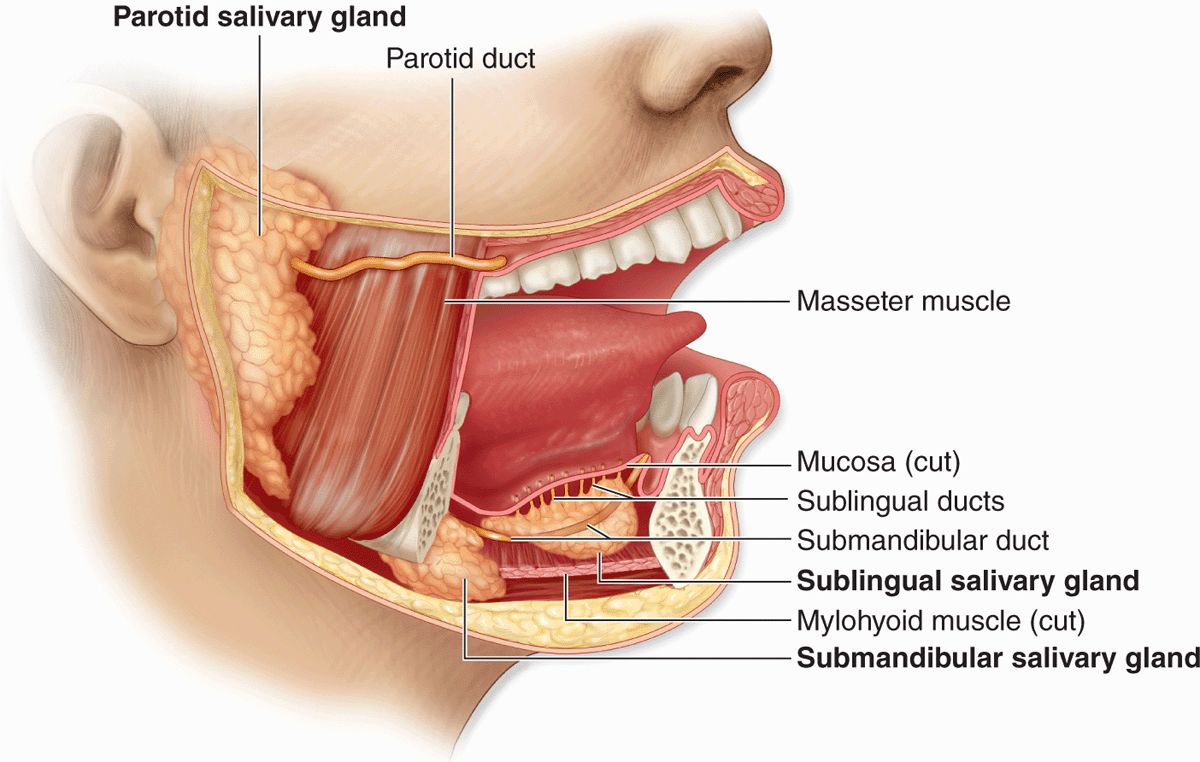
FIGURE 16–1 Major salivary glands.
 MEDICAL APPLICATION
MEDICAL APPLICATION
Inadequate saliva production, leading to dry mouth or xerostomia, can be caused by various factors affecting the major salivary glands, such as mumps viral infection, radiation of the glands, or the normal side effect of drugs such as antihistamines.
A connective tissue capsule surrounds each major salivary gland. The parenchyma of each consists of secretory units on a branching duct system arranged in lobules, separated by septa of connective tissue. The secretion of each gland is either serous, seromucous, or mucous, depending on its content of the glycoprotein mucin. Saliva from the parotids is serous and watery. The submandibular and sublingual glands produce a seromucous secretion, while that of the minor glands is mostly mucous. Saliva is modified by the cells of the duct system draining the secretory units, with much Na+ and Cl– reabsorbed while certain growth factors and digestive enzymes are added.
Three epithelial cell types comprise the salivary secretory units:
 Serous cells are polarized protein-secreting cells, usually pyramidal in shape, with round nuclei, well-stained RER, and apical secretory granules (Figures 16–2 through 16–4). Joined apically by tight and adherent junctions, serous cells form a somewhat spherical unit called an acinus (L. grape), with a very small central lumen (Figure 16–2). Serous acinar cells secrete enzymes and other proteins.
Serous cells are polarized protein-secreting cells, usually pyramidal in shape, with round nuclei, well-stained RER, and apical secretory granules (Figures 16–2 through 16–4). Joined apically by tight and adherent junctions, serous cells form a somewhat spherical unit called an acinus (L. grape), with a very small central lumen (Figure 16–2). Serous acinar cells secrete enzymes and other proteins.
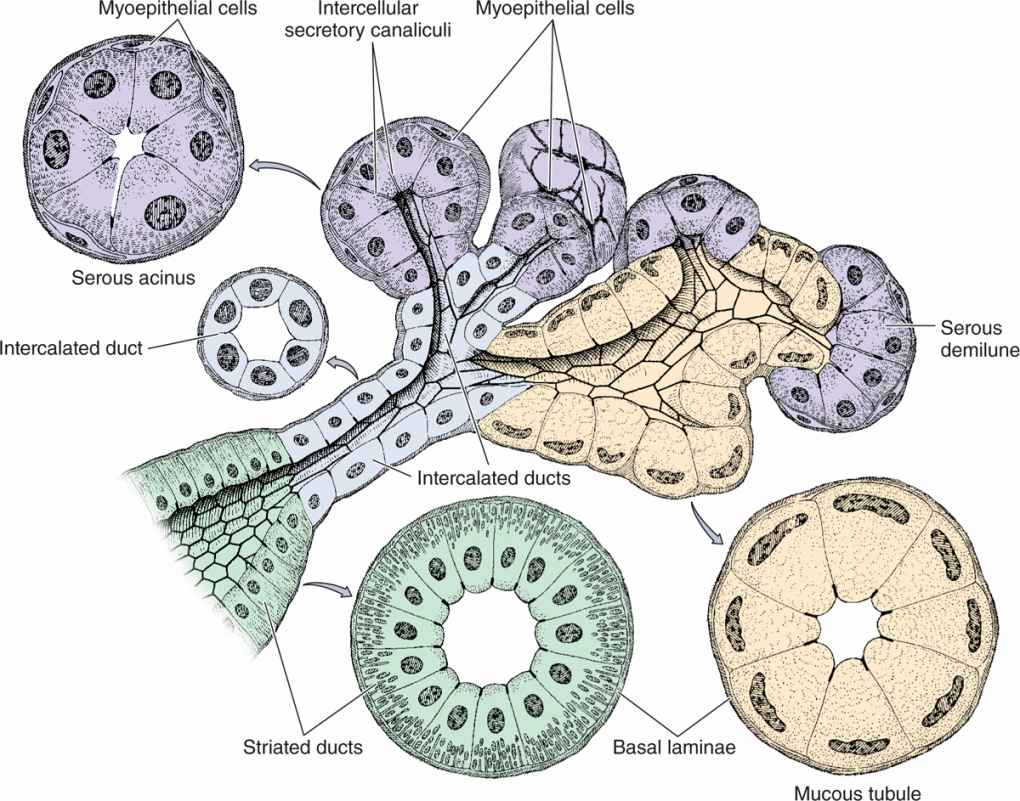
FIGURE 16–2 Epithelial components of a submandibular gland lobule.
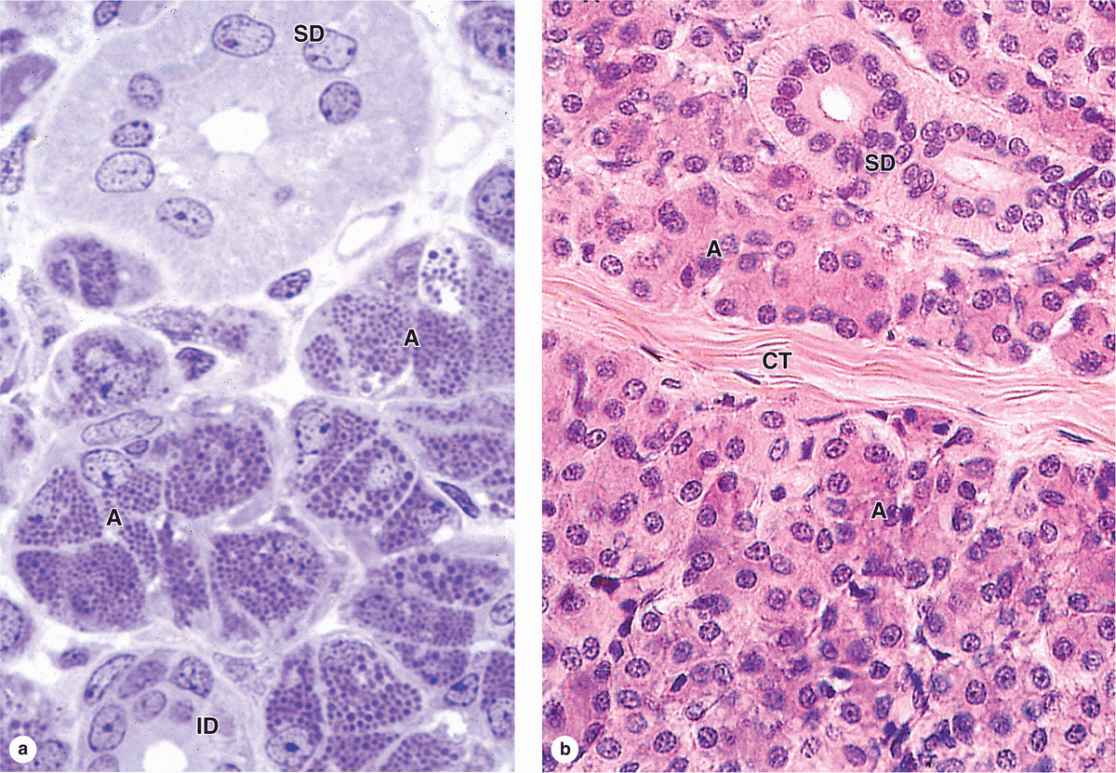
FIGURE 16–3 Parotid gland.
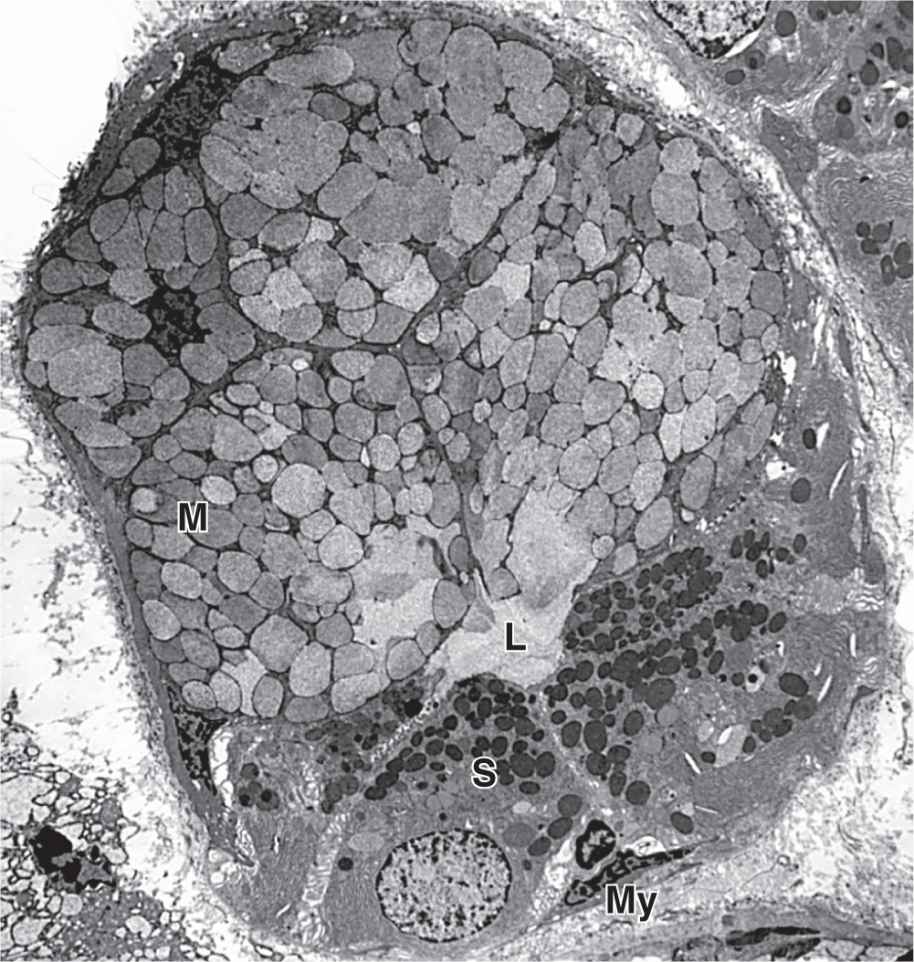
FIGURE 16–4 Ultrastructure of serous and mucous cells.
 Mucous cells are somewhat more columnar in shape, with more compressed basal nuclei (Figures 16–2 and 16–4). Mucous cells contain apical granules with hydrophilic mucins that provide lubricating properties in saliva but cause poor cell staining in routine preparations (Figure 16–5). Mucous cells are most often organized as cylindrical tubules rather than acini. Mixed salivary glands have tubuloacinar secretory units with both serous and mucous secretion.
Mucous cells are somewhat more columnar in shape, with more compressed basal nuclei (Figures 16–2 and 16–4). Mucous cells contain apical granules with hydrophilic mucins that provide lubricating properties in saliva but cause poor cell staining in routine preparations (Figure 16–5). Mucous cells are most often organized as cylindrical tubules rather than acini. Mixed salivary glands have tubuloacinar secretory units with both serous and mucous secretion.
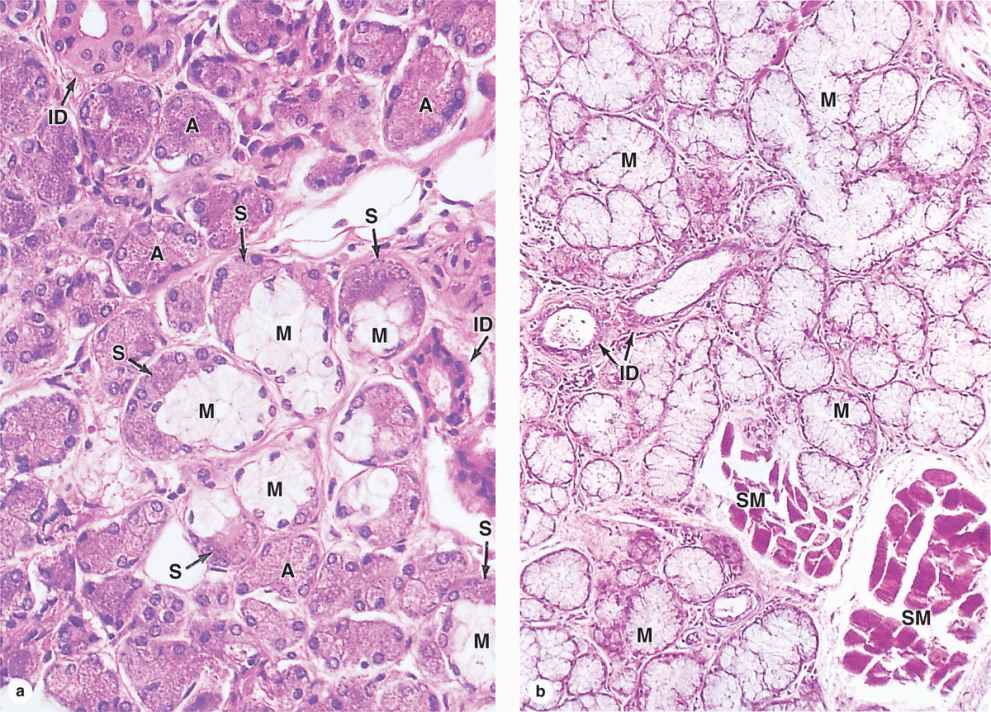
FIGURE 16–5 Submandibular gland and sublingual gland.
 Myoepithelial cells, described in Chapter 4, are found inside the basal lamina surrounding acini, tubules, and the proximal ends of the duct system (Figures 16–2 and 16–4). These small, flattened cells extend several contractile processes around the associated secretory unit or duct and their activity is important for moving secretory products into and through the ducts.
Myoepithelial cells, described in Chapter 4, are found inside the basal lamina surrounding acini, tubules, and the proximal ends of the duct system (Figures 16–2 and 16–4). These small, flattened cells extend several contractile processes around the associated secretory unit or duct and their activity is important for moving secretory products into and through the ducts.

Excessive saliva production, or sialorrhea, is associated with the autonomic activity of nausea, inflammation within the oral cavity, and rabies viral infection.
In the intralobular duct system, secretory acini and tubules empty into short intercalated ducts, lined by cuboidal epithelial cells, and several of these ducts join to form a striated duct (Figure 16–2). The more columnar striated duct cells have many infoldings of their basolateral membrane, all aligned with numerous mitochondria that, by light microscopy, appear as faint basal striations radiating toward the nuclei (Figure 16–6). Striated ducts reabsorb Na+ ions from the initial secretion and their folded cell membranes present a large surface area with ion transporters, facilitating rapid ion transcytosis and making the secretion slightly hypotonic.
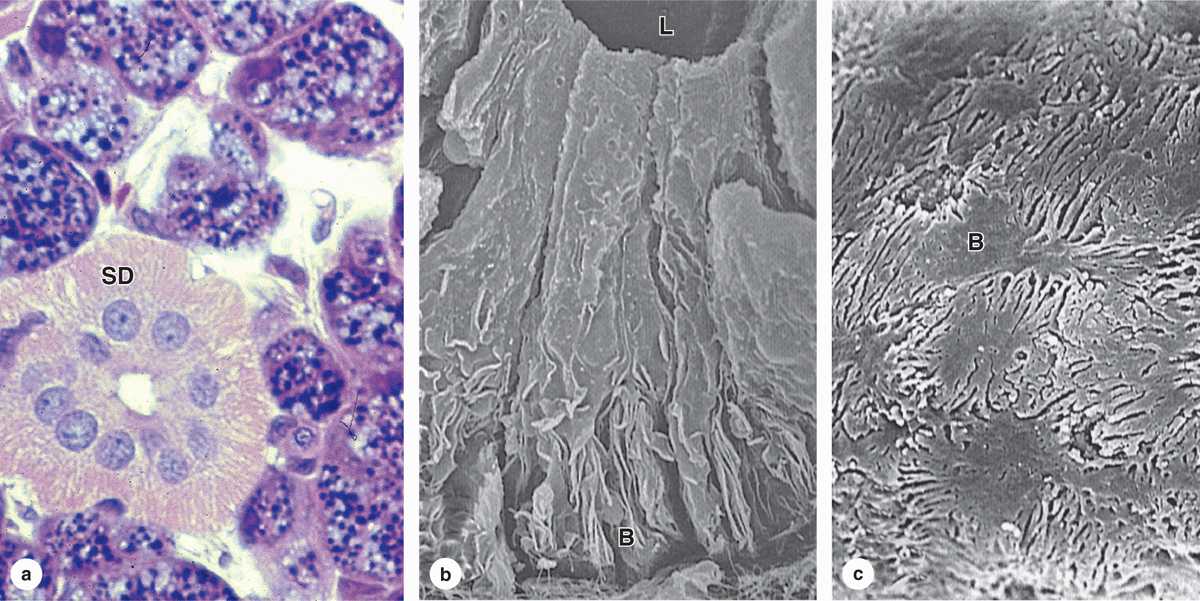
FIGURE 16–6 Striated ducts.
Plasma cells in the connective tissue surrounding the small intralobular ducts release IgA, which forms a complex with the secretory component synthesized by the epithelial cells of the serous acini and intralobular ducts. Transferred into the saliva, the IgA complex released into the saliva provides defense against specific pathogens in the oral cavity.
Ducts from each lobule converge and drain into interlobular excretory ducts with increasing size and thicker connective tissue layers. The lining of these ducts is unusual, combining various epithelial types, including simple cuboidal or columnar, stratified cuboidal or columnar, and pseudostratified epithelia, distributed in no apparent pattern. These atypical epithelia may reflect their composition of cells with many diverse functions, including cells for ion reabsorption, cells for secretion of mucin and other proteins, enteroendocrine cells, and basal stem cells, all in highly branched ducts of small diameter. Before emptying into the oral cavity, the main duct of each gland is lined with nonkeratinized stratified squamous epithelium.
Vessels and nerves enter the large salivary glands at a hilum and gradually branch into the lobules. A rich vascular and nerve plexus surrounds the secretory and duct components of each lobule. The capillaries surrounding the secretory units provide fluid important for saliva production, which is stimulated by the autonomic nervous system. Parasympathetic stimulation, usually elicited through the smell or taste of food, provokes a copious watery secretion with relatively little organic content. Sympathetic stimulation inhibits such secretion and produces the potential for dry mouth often associated with anxiety.
Features specific to each group of major salivary glands include the following:
 Parotid glands, located in each cheek near the ear, are branched acinar glands with exclusively serous acini (Figure 16–3). Serous cells of parotid glands secrete abundant α-amylase that initiates hydrolysis of carbohydrates and proline-rich proteins with antimicrobial and other protective properties.
Parotid glands, located in each cheek near the ear, are branched acinar glands with exclusively serous acini (Figure 16–3). Serous cells of parotid glands secrete abundant α-amylase that initiates hydrolysis of carbohydrates and proline-rich proteins with antimicrobial and other protective properties.
 Submandibular glands, which produce two-thirds of all saliva, are branched tubuloacinar glands, having primarily serous acini, but with many mixed tubuloacinar secretory units (Figures 16–4 and 16-5a). Serous cells of these mixed units are mostly in peripheral crescent-shaped groups called serous demilunes (Figure 16-5a), aggregates formed at least in part artifactually when the adjacent mucous cells swell during routine tissue preparation. Basolateral membrane infoldings of the serous cells facilitate electrolyte and water transport. In addition to a-amylase and proline-rich proteins, serous cells of the submandibular gland secrete other enzymes, including lysozyme for bacterial wall hydrolysis.
Submandibular glands, which produce two-thirds of all saliva, are branched tubuloacinar glands, having primarily serous acini, but with many mixed tubuloacinar secretory units (Figures 16–4 and 16-5a). Serous cells of these mixed units are mostly in peripheral crescent-shaped groups called serous demilunes (Figure 16-5a), aggregates formed at least in part artifactually when the adjacent mucous cells swell during routine tissue preparation. Basolateral membrane infoldings of the serous cells facilitate electrolyte and water transport. In addition to a-amylase and proline-rich proteins, serous cells of the submandibular gland secrete other enzymes, including lysozyme for bacterial wall hydrolysis.
 Sublingual glands, the smallest of the major glands, are also considered branched tubuloacinar glands, but here secretory tubules of mucous cells predominate and the main product of the gland is mucus (Figure 16-6b). The few serous cells present in demilunes add amylase and lysozyme to the secretion.
Sublingual glands, the smallest of the major glands, are also considered branched tubuloacinar glands, but here secretory tubules of mucous cells predominate and the main product of the gland is mucus (Figure 16-6b). The few serous cells present in demilunes add amylase and lysozyme to the secretion.
As described in Chapter 15, small, nonencapsulated salivary glands are distributed throughout the oral mucosa and submucosa with short ducts to the oral cavity. These minor salivary glands are usually mucous, except for the small serous glands at the bases of circumvallate papillae. Plasma cells releasing IgA are also common within the minor salivary glands.
PANCREAS
The pancreas is a mixed exocrine-endocrine gland that produces both digestive enzymes and hormones. It is an elongated retroperitoneal organ, with a large head near the duodenum and more narrow body and tail regions that extend to the left (Figure 16–7). The pancreas has a thin capsule of connective tissue, from which septa extend to cover the larger vessels and ducts and to separate the parenchyma into lobules (Figure 16–8). The secretory acini are surrounded by a basal lamina that is supported only by a delicate sheath of reticular fibers with a rich capillary network. Endocrine function of the pancreas involves primarily smaller cells similar to enteroendocrine cells located in variously sized clusters called the pancreatic islets (islets of Langerhans). These are described with the endocrine organs in Chapter 20.
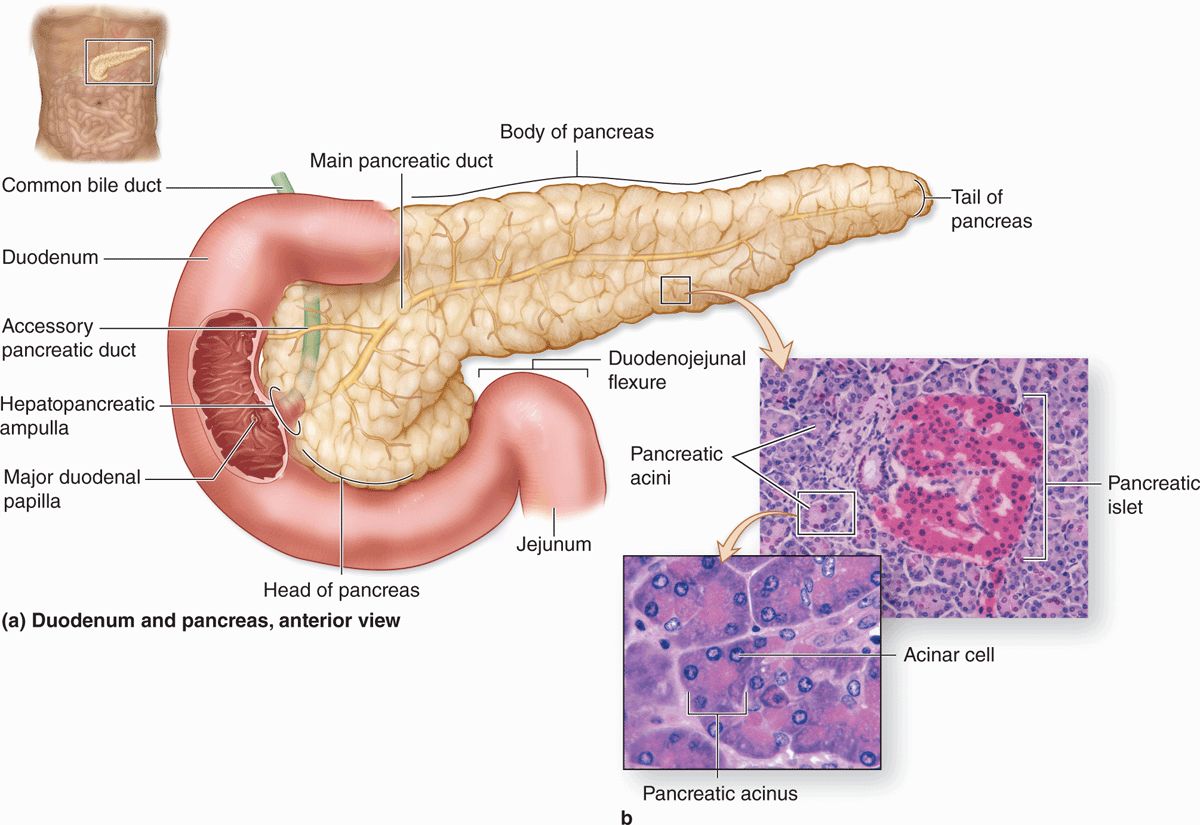
FIGURE 16–7 Pancreas and duodenum.
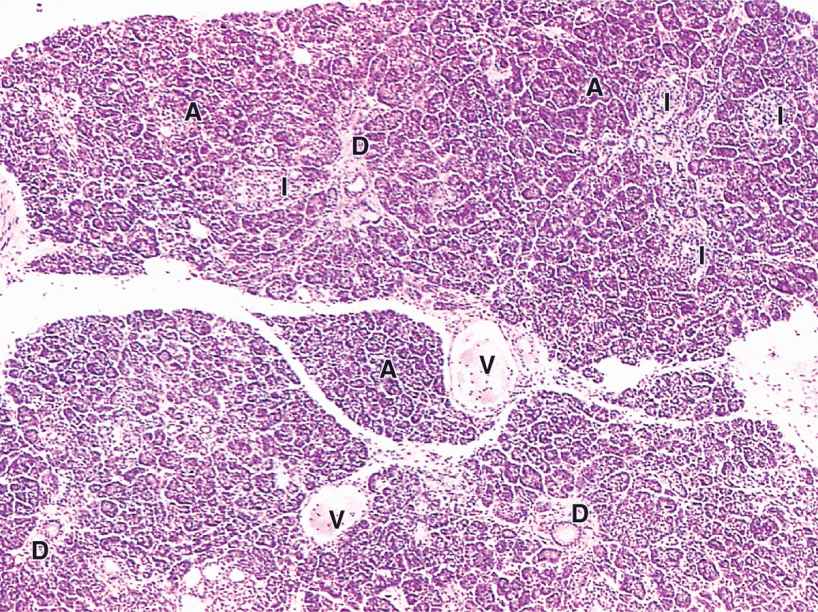
FIGURE 16–8 Pancreas.

Pancreatic cancer, which is usually a carcinoma of duct cells, can arise anywhere in the gland but occurs most often in the head of the organ near the duodenum. The tumor is usually asymptomatic until growth and metastasis are well advanced, leading to the low rate of early detection and subsequent high rate of mortality. Metastasis may be facilitated by the relatively sparse connective tissue around the ducts and vasculature of the pancreas.
The digestive enzymes are produced by cells of serous acini in the larger exocrine portion of the pancreas (Figure 16-9a). This somewhat resembles the parotid gland histologically, although the pancreas lacks striated ducts and the parotid glands lack islets of endocrine tissue. Each pancreatic acinus consists of several serous cells surrounding a very small lumen, without myoepithelial cells (Figure 16–9). The acinar cells are polarized, with round basal nuclei, and numerous zymogen granules apically, typical of protein-secreting cells (Figure 16–10).
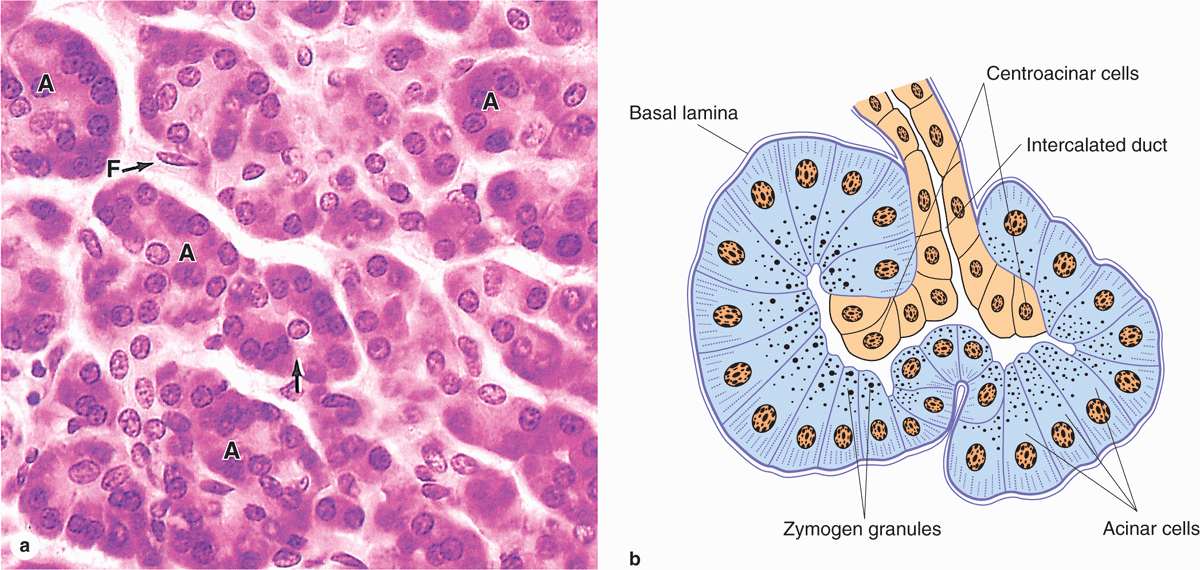
FIGURE 16–9 Pancreatic acini.
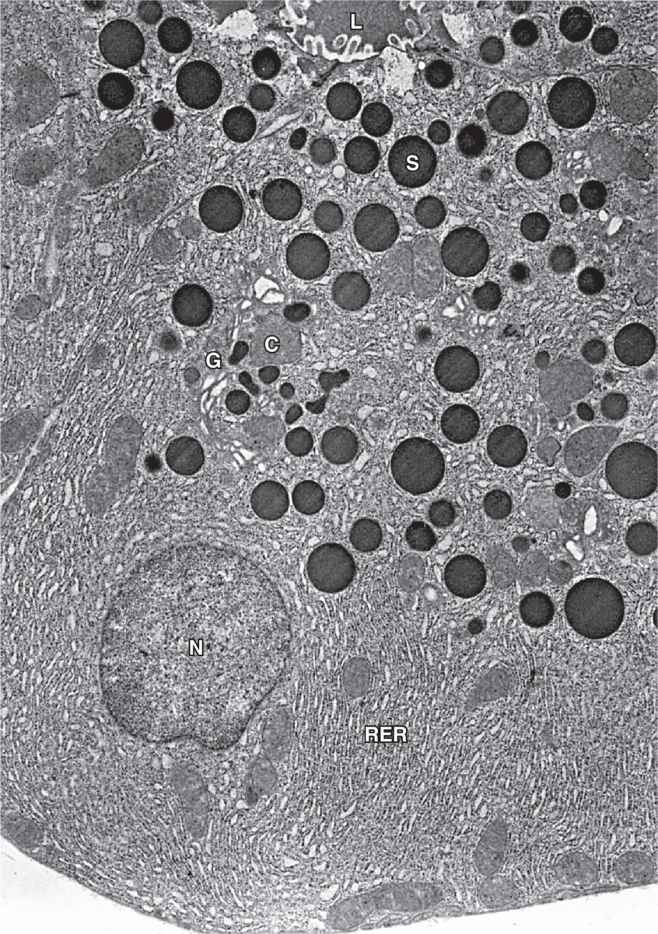
FIGURE 16–10 Pancreatic acinar cell ultrastructure.
Each acinus is drained by a short intercalated duct of simple squamous epithelium. The initial cells of these small ducts extend into the lumen of the acinus as small pale-staining centroacinar cells that are unique to the pancreas. Cells of the intercalated ducts secrete a large volume of fluid, rich in HCO3– (bicarbonate ions), which alkalinizes and transports hydrolytic enzymes produced in the acini. The intercalated ducts merge with intralobular ducts and larger interlobular ducts, which have increasingly columnar epithelia before joining the main pancreatic duct that runs the length of the gland.
The exocrine pancreas secretes approximately 1.5 L of alkaline pancreatic juice per day and delivers it directly into the duodenum where the HCO3– ions neutralize the acidic chyme entering there from the stomach and establish the pH for optimal activity of the pancreatic enzymes. These digestive enzymes include several proteases, α-amylase, lipases, and nucleases (DNAase and RNAase). The proteases are secreted as inactive zymogens (trypsinogen, chymotrypsinogen, proelastase, kallikreinogen, and procarboxipeptidases). Trypsinogen is cleaved and activated by enteropeptidases in the duodenum, generating trypsin that activates the other proteases in a cascade. Pancreatic tissue is protected against autodigestion by the following:
 Restricting protease activation to the duodenum,
Restricting protease activation to the duodenum,
 Trypsin inhibitor, which is copackaged in the secretory granules with trypsinogen, and
Trypsin inhibitor, which is copackaged in the secretory granules with trypsinogen, and
 The low pH in the acini and duct system due to HCO3–secreted by the centroacinar and intercalated duct cells, which helps keep all the enzymes inactive.
The low pH in the acini and duct system due to HCO3–secreted by the centroacinar and intercalated duct cells, which helps keep all the enzymes inactive.



