CHAPTER 11 Membrane Physiology
This chapter describes how pumps, carriers, and channels cooperate in living systems. These three components often work together in circuits or cycles. Pumps establish gradients of ions across membranes (see Chapter 8). Channels regulate membrane permeability to these ions to maintain the electrical potential (see Chapter 10) required for membrane excitability. Carriers use ion gradients as a source of energy to drive transport as well as to do other work (see Chapter 9). Coupling ion fluxes through pumps and carriers to do work is called a chemiosmotic cycle.
Chemiosmotic Cycles
A simple chemiosmotic cycle couples a cation transporting pump to solute transport by a carrier (Fig. 11-1). The membrane could be a plasma membrane or an organelle membrane. The driving reaction is called the primary transport step, indicating an input of energy and, in most cases, some chemical reaction. Other reactions are called secondary transport reactions, indicating that they depend on ion gradients. The transported substrate is the same chemically on both sides of the mem-brane. Although they are simple in concept, the importance and power of chemiosmotic cycles should not be underestimated. They operate in every membrane of every cell.
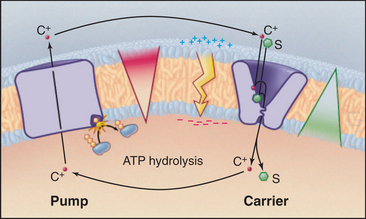
Pumps use energy derived from ATP hydrolysis, light absorption, or another chemical reaction (see Table 8-1) to move ions in one direction across a membrane. This raises the concentration of a cation (C+) on one side and depletes it on the other side of a membrane-bounded compartment. An ion gradient is characterized by both a chemical term, the concentration gradient, and an electrical gradient (the membrane potential explained in Fig. 10-17). The electro-chemical potential across a membrane represents a reservoir of power and a capacity to do work, also known as an ion-motive force. A mechanical analog would be using a pump to fill an elevated reservoir with fluid.
Carriers and other membrane proteins use the potential energy of ion gradients to drive other processes. This is analogous to using fluid flow out of a reservoir to drive a turbine, which puts the energy to use for other types of work. Many carriers use energy derived from the downhill passage of one substrate to transport one or more other substances up their concentration gradients across the same membrane barrier. In Figure 11-1, the carrier links the transport of solute S to the movement of cation C+ down its gradient. Recirculation of cations allows a cell to accumulate solute against its concentration gradient. In addition to the osmotic work illustrated in the figure, chemiosmotic cycles can do chemical work. During both oxidative and photosynthetic phosphorylation, proton cycles drive ATP synthesis by F0F1-ATP synthases (see Fig. 19-5). Chemiosmotic cycles can also perform mechanical work. The electrochemical gradient of protons across the plasma membrane drives rotation of bacterial flagella (see Fig. 38-24).
Epithelial Transport
Net transport across an epithelium depends on tight junctions (see Fig. 31-2) that seal the extracellular space between the cells (Fig. 11-2). These junctions separate two extracellular compartments. The apical compartment is the free surface (e.g., the skin) or the lumen of the organ (e.g., the intestine, respiratory tract, or kidney tubules). The basolateral compartment lies between epithelial cells and is continuous with the underlying connective tissue and its blood vessels. Tight junctions seal the extracellular space, inhibiting diffusion of solutes, between the apical and basolateral compartments of the extracellular space. The extent of this seal varies from very tight to leaky. Tight junctions also separate the plasma membrane into apical and basolateral domains, restricting the movement of integral membrane proteins between these domains.
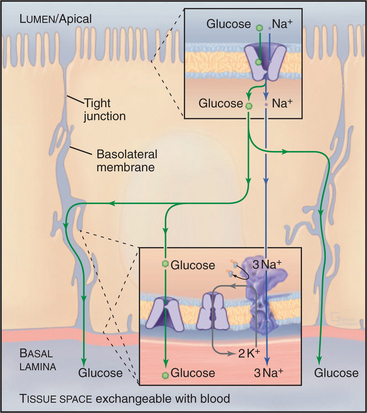
Glucose Transport in the Intestine, Kidney, Fat, and Muscle
A chemiosmotic cycle transports glucose uphill from the lumen of the intestine to the blood (Fig. 11-2). Tight junctions restrict movement of glucose between the epithelial cells, so all of the glucose must move through the cytoplasm. Glucose transport across the epithelial cells requires the following components:
Glucose uptake by fat and muscle cells offers a different perspective. These tissues are designed to take up glucose from the blood when it is plentiful following a meal. Mammals have genes for six isoforms of the classical D-glucose uniporter. Insulin in the blood regulates the availability of the GLUT4 isoform in the plasma membrane. Muscle and fat express GLUT4 but store it internally in membrane vesicles. After a meal, high blood glucose stimulates secretion of insulin into blood. Signal transduction mechanisms (see Fig. 27-7) lead to fusion of these GLUT4 vesicles with the plasma membrane. That increases the rate of glucose transport into fat and muscle by 5-fold to 20-fold, lowering the blood glucose concentration and providing these cells with glucose, which they then convert to glycogen and triglycerides for storage.
Salt and Water Transport in the Kidney
In a section of the kidney tubule called the loop of Henle, the epithelium uses Na+K+-ATPase pumps and Na+/K+/2Cl− symporters to reabsorb NaCl that is filtered from blood into the excretory pathway (Fig. 11-3). Without this provision, salt would be lost in urine. Tight junctions seal this epithelium, so that salt must pass through the cells to return to the blood. Na+/K+/2Cl− symporters in the apical plasma membrane provide a way for NaCl to enter the cell down its concentration gradient. Abundant Na+K+-ATPases in the basolateral plasma membrane (5000/mm2) create a Na+ gradient to drive the symporter and to clear the cytoplasm of Na+ accumulated from the tubule lumen. KCl that enters with Na+ through the Na+/K+/2Cl− symporter leaves the cell through channels: K+ channels in apical and basolateral membranes and Cl− channels in basolateral membranes.
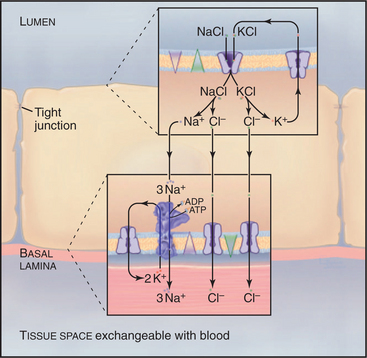
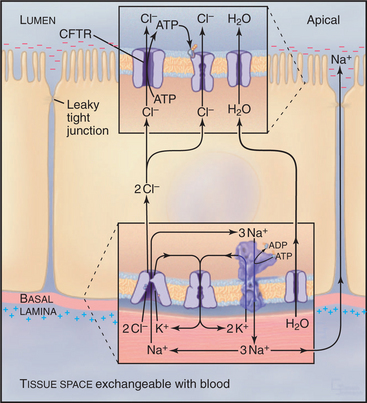
Cystic Fibrosis as a Transporter Disease
Normally, cells in the lung and gastrointestinal tract use a complicated selection of familiar pumps and carriers to secrete salt and water at their apical sur-faces (Fig. 11-4). Na+K+-ATPases in the basolateral membrane set up an electrochemical gradient of Na+, which is exploited by basolateral membrane Na+/K+/2Cl− symporters to take in Na+, along with K+ and Cl− anions. The inward movement of Na+ down its electrochemical gradient drives the entry of K+ and Cl− up their gradients. This brings excess potassium chloride into the cell. (The K+ brought in by both the Na+K+-ATPase and the Na+/K+/2Cl− symporter recycles after exiting from the cell by way of channels in the basolateral plasma membrane. Thus, K+ is merely catalytic.) Excess Cl− is left inside the cell. The cystic fibrosis transmembrane regulator (CFTR) protein, an ABC pump, in the apical plasma membrane acts as a Cl− channel. When protein kinase phosphorylates the regulatory domain and ATP binds to the cytoplasmic domains, a conformational change opens a Cl− channel across the membrane. Cl− moves down its electrochemical gradient out of the cell, carrying charge to the outside. The whole epithelium becomes polarized, with the lumen electrically negative relative to the extracellular fluid compartment. This electrical driving force allows Na+ to move between cells, from the extracellular fluid compartment through leaky tight junctions to the surface of the epithelium. Sodium chloride on the apical surface creates an osmotic force that draws water down its concentration gradient across the cells to the outside through water channels (see Fig. 10-15). CFTR also appears to inhibit the transport mechanisms that reabsorb fluid from the lumen of the epithelium. A balance between this fluid secretion and fluid reabsorption normally keeps the surface of the epithelium properly hydrated, allowing the cilia to clear the lung of bacteria and secretions and the ducts of the pancreas to secrete digestive enzymes.
Cellular Volume Regulation
Cells employ both short- and long-term strategies involving pumps, carriers, and channels to maintain a constant volume (Fig. 11-5). These compensatory mechanisms are required because water moves across the plasma membrane through water channels and slowly through lipid bilayers if the osmotic strength of the environment differs even slightly from that inside the cell. Water moves to maintain an osmotic equilibrium, as is illustrated for red blood cells in Figure 7-6. In a hypotonic medium, water moves into a cell to dilute the cytoplasm. In a hypertonic medium, water moves out to concentrate the cytoplasm. Mechanisms that are employed to compensate for these volume changes are well defined, but the mechanisms that sense volume changes and trigger these responses are still being investigated.
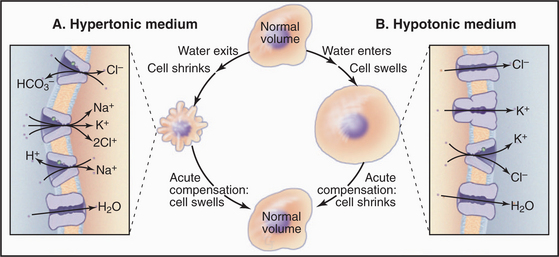
Animal cells respond acutely to loss of water by activating Na+/H+ antiporters, Cl−/HCO3− antiporters, and/or Na+/K+/2Cl− symporters that bring potassium chloride and sodium chloride into the cell. Water follows, returning the cell to its original volume in minutes. The acute response to swelling activates K+ channels, Cl− channels (ClC-3), and/or a K+/Cl− symporter, taking potassium chloride and water out of the cell. Compensation by moving inorganic ions works in the short run but is not an acceptable long-term solution because changes in the internal concentrations of K+, Na+, and Cl− affect the membrane potential and other physiological processes.
Excitable Membranes
Electrical excitability is not limited to nerves and muscles. Eggs use a form of action potential as an early step in blocking fertilization by more than one sperm. Chemotaxis by macrophages and secretion of insulin and other hormones both depend on electrical excitability. The reader should be familiar with the appendixes in Chapter 10 to appreciate the following material.
Description of an Action Potential
If a microelectrode (see Fig. 10-16A) drives a small positive or negative current into a cell, a second microelectrode a short distance away detects a small voltage response. These electrotonic potentials decline rapidly with distance if the cell in nonexcitable.
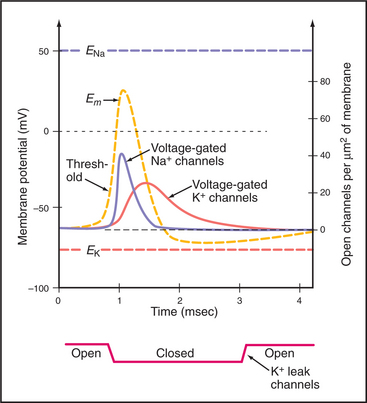
In striking contrast to these small local currents, when the plasma membrane of an excitable cell, such as neuron or muscle, is depolarized beyond a certain level, called a threshold, the membrane responds over a few milliseconds with a large, stereotyped change in membrane potential, called an action potential (Fig. 11-6). Voltage-gated ion channels (see Fig. 10-7) generate this powerful electrical signal that spreads rapidly (10 m/sec) over the entire plasma membrane. During an action potential, the membrane potential can reach a peak of +40 to 50 mV before repolarizing to the resting potential. Because action potentials are self-triggering, they travel without dissipation over long distances. This high-speed transmission is very efficient, requiring movement of very few ions across the membrane.