primarily as MgATP2-. Mg therefore is essential for the function of the glycolytic cycle, citric acid cycle, protein kinases, RNA and DNA polymerases, lipid metabolism, and amino acid activation, as well as playing a critical role in the cyclic adenosine monophosphate (cAMP) and phospholipase C second-messenger systems (5, 6, 10, 11, 12).
TABLE 9.1 DISTRIBUTION AND CONCENTRATIONS OF MAGNESIUM (Mg) IN A HEALTHY ADULT (TOTAL BODY: 833-1170 mmola, OR 20-28 g) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
of Mg absorption and may be influenced by certain hormones (39). Absorption of Mg as a function of intake is curvilinear (Fig. 9.3), and this pattern reflects this active saturable process and passive diffusion. Net Mg absorption increases with increasing Mg intake; however, fractional Mg absorption falls. When small amounts of Mg were fed in the form of a standard meal supplemented by varying amounts of Mg (40), fractional absorption fell progressively from approximately 65% to 70% with intake of 7 to 36 mg (0.3 to 1.5 mmol) down to 11% to 14% with intake of 960 to 1000 mg (40 mmol).
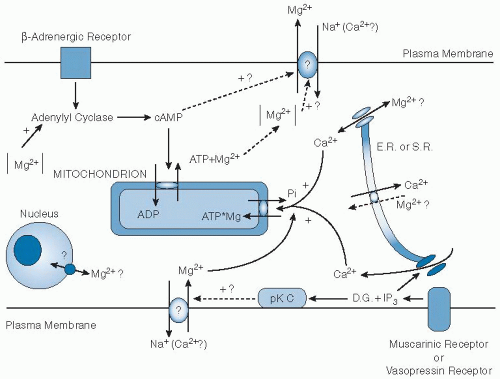 Fig. 9.1. Schema of regulation of cellular magnesium (Mg2+) homeostasis in the mammalian cell. The pathways are indicated for cellular Mg2+ release (upper section) and for its uptake (lower section). Stimulated by β-adrenergic agonists, cyclic adenosine monophosphate (cAMP) is increased in the cytosol, which modulates mitochondrial adenine nucleotide translocase and increases the efflux of Mg2+ from the mitochondrion by means of an exchange of one Mg adenosine triphosphate (MgATP) for adenosine diphosphate (ADP). Activation of muscarinic receptors (in cardiac cells) or vasopressin receptors (in the liver) may stimulate an Mg2+ influx mechanism either by decreasing cAMP or by enhancing protein kinase C (pK C) activity by diacylglycerol (D.G.). Vasopressin receptor activation is coupled with production of inositol triphosphate (IP3) from phosphatidylinositol bisphosphate, which induces release of calcium (Ca2+) from the endoplasmic reticulum (E.R.) or the sarcoplasmic reticulum (S.R.). Ca2+ release may be associated with either Mg2+ influx or Mg redistribution in the nucleus or endoplasmic reticulum. Na+, sodium. (Adapted with permission from Romani A, Marfella C, Scarpa A. Cell magnesium transport and homeostasis: role of intracellular compartments. Miner Electrolyte Metab 1993;19:282-9.) |
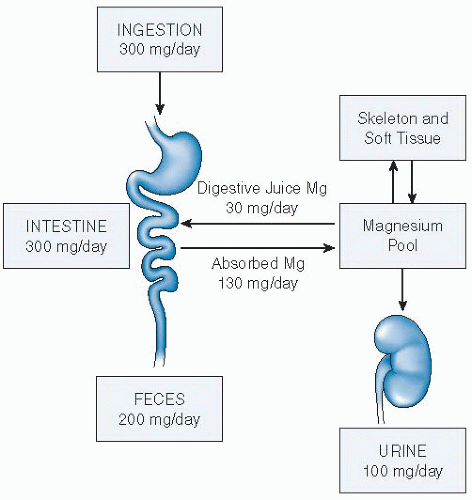 Fig. 9.2. Magnesium (Mg) homeostasis in humans. A schematic representation of its metabolic economy indicating (a) its absorption from the alimentary tract, (b) its distribution into bone, and (c) its dependence on the kidney for excretion. Homeostasis depends on the integrity of intestinal and renal absorptive processes. (Adapted with permission from Rude RK. Magnesium homeostasis. In: Bilezikian JB, Raisz L, Rodan G, eds. Principles of Bone Biology. 3rd ed. San Diego: Academic Press, 2008:487-513.) |
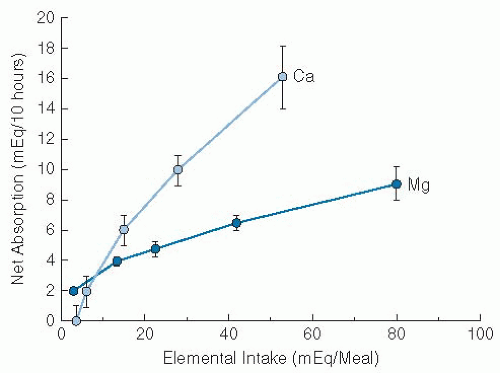 Fig. 9.3. Net magnesium (Mg) and calcium (Ca) absorption in healthy humans. The data were obtained under conditions described in reference 39 and in the text. Mean values S.E. are indicated by vertical bars. The absorption data for Mg represent a curved function compatible with a saturable process (at ˜10 mEq/meal in this study) and a linear function reflecting passive diffusion at higher intakes. (Adapted with permission from Fine KD, Santa Ana CA, Porter JL et al. Intestinal absorption of magnesium from foods and supplements. J Clin Invest 1991;88:396-402.) |
the nephron is summarized in Figure 9.4. Paracellin-1 (claudin-16) and claudin-19 appear to mediate this transport (27, 28). The distal convoluted tubule reabsorbs 5% to 10% of filtered Mg via an active transcellular pathway. Several proteins may be involved, including the sodium chloride cotransporter (28). TRPM6 is also expressed in the distal tubule. Mutations of TRPM6 result in lower intestinal Mg absorption and renal Mg wasting (27, 28, 29).
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


