16 LOWER DIGESTIVE SEGMENT
SMALL INTESTINE
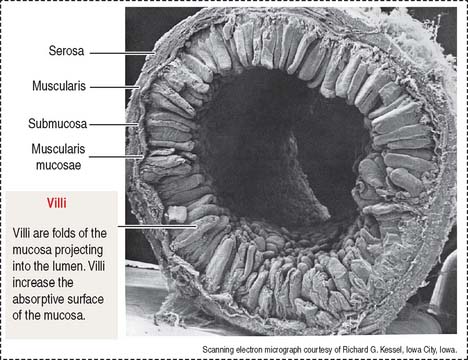
The wall of the small intestine consists of four layers (Figures 16-1 to 16-3): (1) the mucosa, (2) the submucosa, (3) the muscularis, and (4) the serosa, or peritoneum. As you will see, histologic differences are seen in the mucosa and submucosa of the three major portions of the small intestine. The muscularis externa and serosa layers are similar.
Intestinal wall
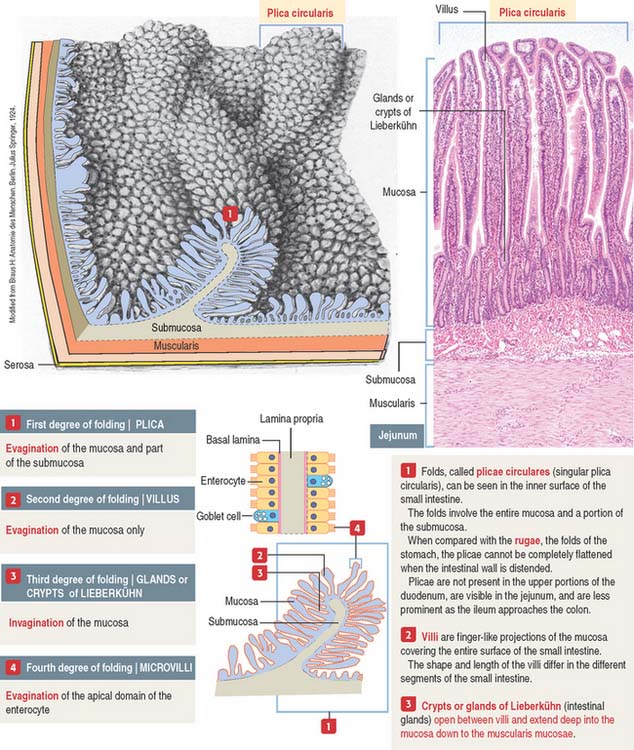
An increase in the total surface of the mucosa reflects the absorptive function of the small intestine. Four degrees of folding amplify the absorptive surface area of the mucosa (see Figure 16-2): (1) the plicae circulares (circular folds; also known as the valves of Kerkring), (2) the intestinal villi, (3) the intestinal glands, and (4) the microvilli on the apical surface of the lining epithelium of the intestinal cells (enterocytes).
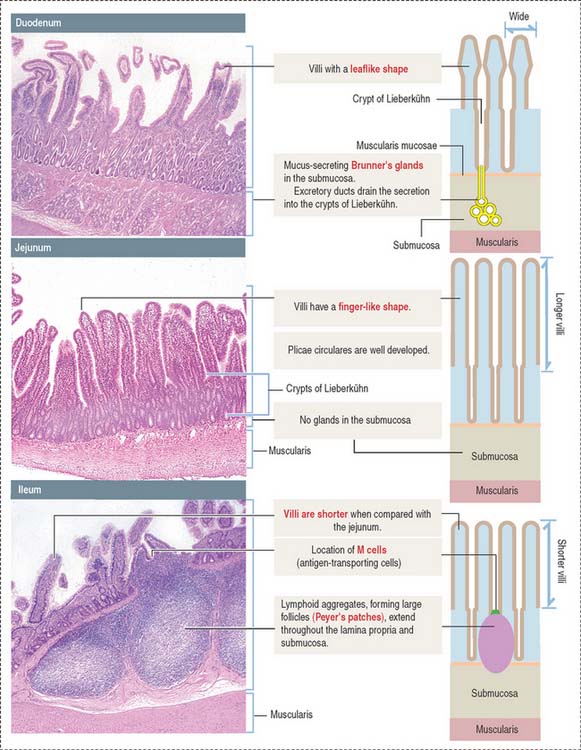
A plica circularis is a permanent fold of the mucosa and submucosa encircling the intestinal lumen.
The intestinal villi are finger-like projections of the mucosa covering the entire surface of the small intestine. Villi extend deep into the mucosa to form crypts ending at the muscularis mucosae. The length of the villi depends on the degree of distention of the intestinal wall and the contraction of smooth muscle fibers in the villus core.
The muscularis mucosae is the boundary between the mucosa and submucosa (see Figure 16-3). The muscularis consists of inner circular smooth muscle and outer longitudinal smooth muscle. The muscularis is responsible for segmentation and peristaltic movement of the contents of the small intestine (Figure 16-4). A thin layer of loose connective tissue is covered by the visceral peritoneum, a serosal layer lined by a simple squamous epithelium, or mesothelium. The parietal peritoneum covers the inner surface of the abdominal wall.
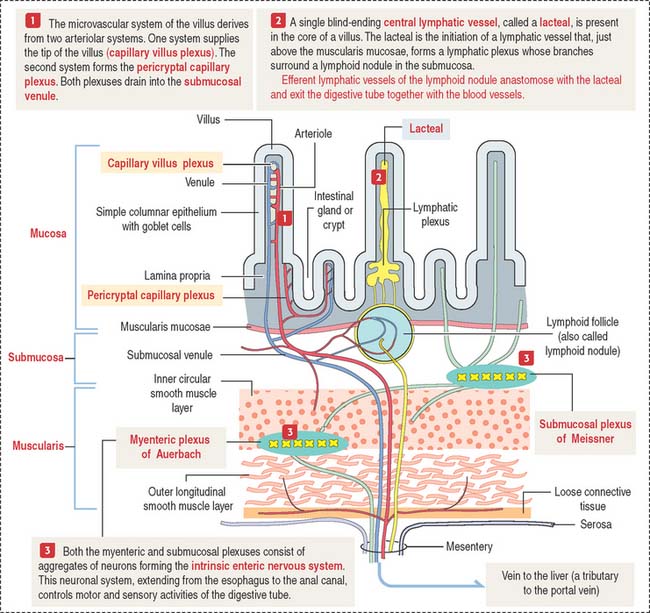
Microcirculation of the small intestine
A difference from the microcirculation of the stomach (see Figure 15-8 in Chapter 15, Upper Digestive Segment) is that the intestinal submucosa is the main distribution site of blood and lymphatic flow (see Figure 16-3).
Innervation and motility of the small intestine
Motility of the small intestine is controlled by the autonomic nervous system. The intrinsic autonomic nervous system of the small intestine, consisting of the submucosal plexus of Meissner and myenteric plexus of Auerbach, is similar to that of the stomach (see Figure 15-9 in Chapter 15, Upper Digestive Segment).
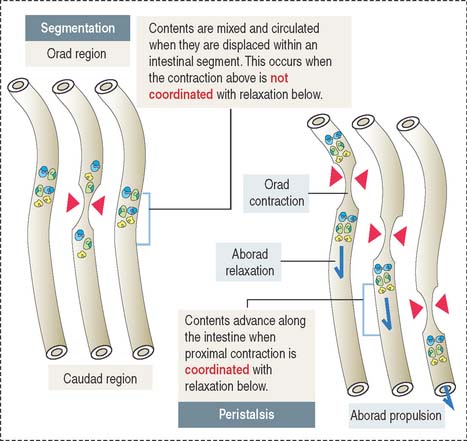
Contraction of the muscularis is coordinated to achieve two objectives (see Figure 16-4): (1) To mix and mobilize the contents within an intestinal segment. This is accomplished when muscular contraction activity is not coordinated and the intestine becomes transiently divided into segments. This process is known as segmentation. (2) To propel the intestinal contents when there is a proximal (orad) contraction coordinated with a distal (aborad; Latin ab, from; os, mouth; away from the mouth) relaxation. When coordinated contraction-relaxation occurs sequentially, the intestinal contents are propelled in an aborad direction. This process is known as peristalsis (Greek peri, around; stalsis, constriction).
Histologic differences between the duodenum, jejunum, and ileum
Each of the three major anatomic portions of the small intestine—the duodenum, jejunum, and ileum—has distinctive features that allow recognition under the light microscope (Figure 16-5).
Villi and crypts of Lieberkühn
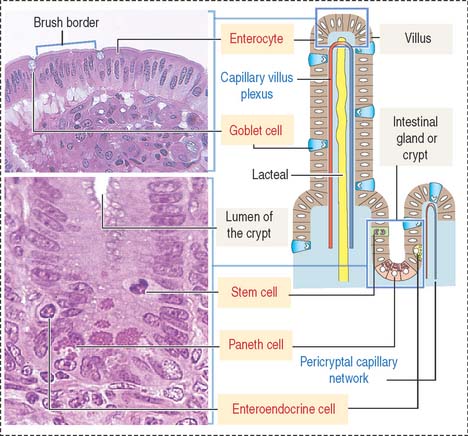
The intestinal mucosa, including the crypts of Lieberkühn, are lined by a simple columnar epithelium containing four major cell types (Figure 16-6): (1) absorptive cells, or enterocytes, (2) goblet cells, (3) Paneth cells, and (4) enteroendocrine cells. Stem cells, Paneth cells, and enteroendocrine cells are found in the crypts of Lieberkühn (see Figure 16-6).
Absorptive intestinal cells, or enterocytes
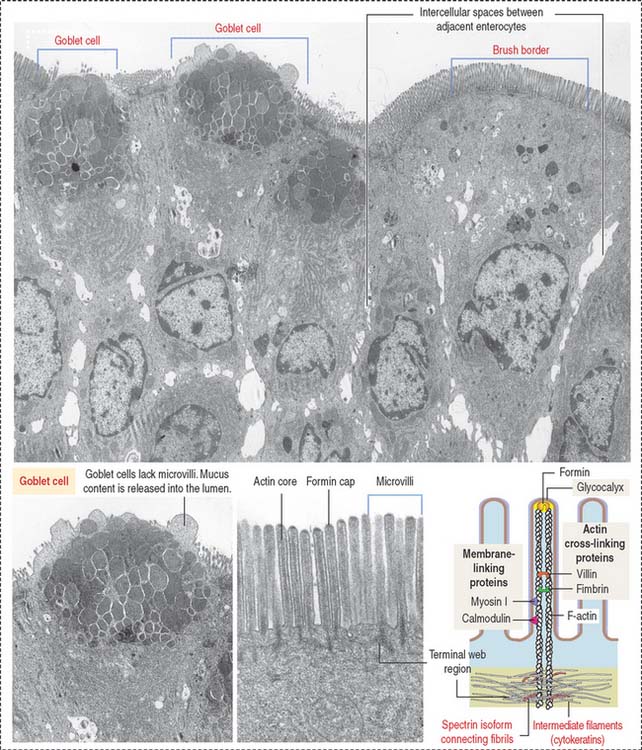
The absorptive intestinal cell or enterocyte has an apical domain with a prominent brush border (also called a striated border), ending on a clear zone, called the terminal web, which contains transverse cytoskeletal filaments. The brush border of each absorptive cell contains about 3000 closely packed microvilli, which increase the surface luminal area 30-fold.
The length of a microvillus ranges from 0.5 to 1.0 μm. The core of a microvillus (Figure 16-7) contains a bundle of 20 to 40 parallel actin filaments cross-linked by fimbrin and villin. The actin bundle core is anchored to the plasma membrane by formin (protein of the cap), myosin I, and the calcium-binding protein calmodulin. Each actin bundle projects into the apical portion of the cell as a rootlet, which is cross-linked by an intestinal isoform of spectrin to an adjacent rootlet. The end portion of the rootlet attaches to cytokeratin-containing intermediate filaments. Spectrin and cytokeratins form the terminal web. The terminal web is responsible for maintaining the upright position and shape of the microvillus and anchoring the actin rootlets.
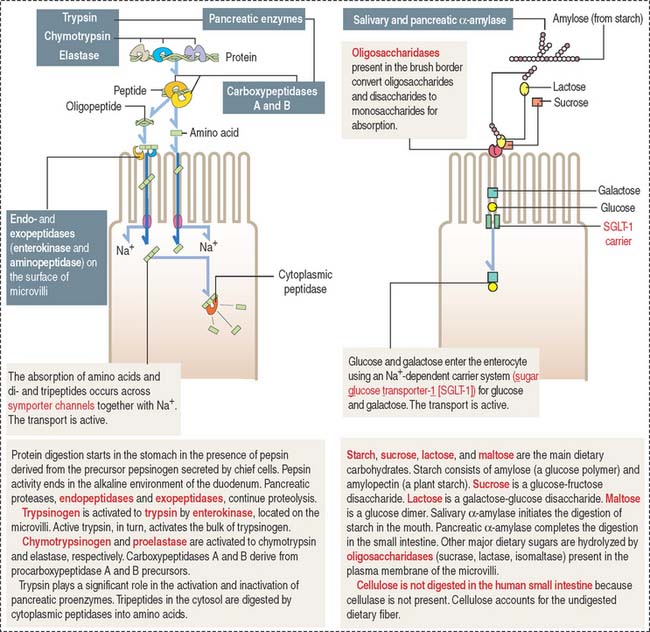
The microvilli, forming a brush border, contain intramembranous enzymes, including lactase, maltase, and sucrase (Figure 16-8). These oligosaccharides reduce carbohydrates to hexoses, which can be transported into the enterocyte by carrier proteins. A genetic defect in lactase prevents the absorption of lactose-rich milk, leading to diarrhea (lactose intolerance). Therefore, the brush border not only increases the absorptive surface of enterocytes but is also the site where enzymes are involved in the terminal digestion of carbohydrates and proteins.
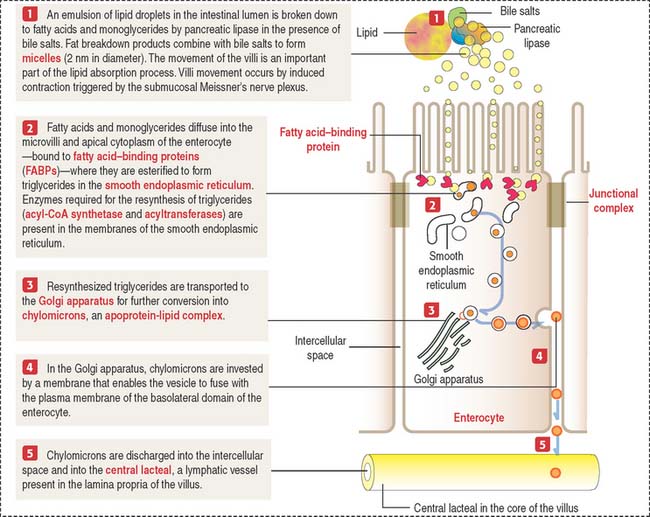
The absorption of lipids involves the enzymatic breakdown of dietary lipids into fatty acids and monoglycerides, which can diffuse across the plasma membrane of the microvilli and the apical plasma membrane of the enterocyte. Details of the process of fat absorption are depicted in Figure 16-9.
Goblet cells
Goblet cells are columnar mucus-secreting cells scattered among enterocytes of the intestinal epithelium (see Figure 16-7).
Goblet cells have two domains: (1) a cup- or goblet-shaped apical domain containing large mucus granules that are discharged on the surface of the epithelium and (2) a narrow basal domain, which attaches to the basal lamina and contains the rough endoplasmic reticulum in which the protein portion of mucus is produced. The Golgi apparatus, which adds oligosaccharide groups to mucus, is prominent and situated above the basally located nucleus.
Enteroendocrine cells
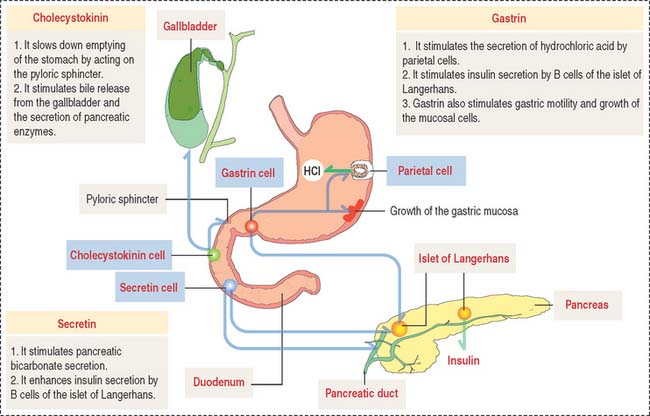
We have already studied the structural and functional features of enteroendocrine cells in the stomach (see Chapter 15, Upper Digestive Segment). As in the stomach, enteroendocrine cells secrete peptide hormones controlling several functions of the gastrointestinal system. The location and function of gastrin-, secretin-, and cholecystokinin-secreting cells are summarized in Figure 16-10.
PROTECTION OF THE SMALL INTESTINE
The large surface area of the gastrointestinal tract, about 200 m2 in humans, is vulnerable to resident microorganisms, called microbiota, and potentially harmful microorganisms and dietary antigens. We discussed in Chapter 15, Upper Digestive Segment, the role of the mucus blanket in the protection of the surface of the stomach during Helicobacter pylori infection. In the small and large intestines, goblet cells secrete mucin glycoproteins assembled into a viscous gel-like blanket limiting direct bacterial contact with enterocytes. A lack of mucin glycoprotein 2 (MIC2) causes spontaneous intestinal inflammation.



