CHAPTER 37 Intracellular Motility
Virtually every component inside living cells moves to some extent, but the magnitude and velocity of these movements vary by orders of magnitude depending on the cell (Table 37-1 and Fig. 37-1). At one extreme, the bulk cytoplasm of algae and giant amoebas streams tens of micrometers per second. At the other extreme, small molecules and macromolecules diffuse through cytoplasm essentially unnoticed. The network of cytoskeletal polymers has a pore size of less than 50 nm, so particles that are larger than the pores must be transported actively. For example, messenger RNA (mRNA) moves from its site of synthesis in the nucleus through nuclear pores into the cytoplasm and then may be carried actively to specific parts of the cell. The nucleus rotates back and forth in most cells. Lysosomes, mitochondria, secretory vesicles, and endosomes all move around actively in cytoplasm, frequently between the centrosome and the cell periphery. Intracellular pathogenic bacteria and viruses subvert the host cell’s actin system to propel themselves randomly through the cytoplasm. Virus particles move along microtubules.
Table 37-1 VELOCITIES OF INTRACELLULAR MOVEMENTS
| System | Velocity (μm s−-1) | Mechanism |
|---|---|---|
| Microtubule Motors | ||
| Anterograde fast axonal transport, squid | 1 | Individual kinesin motors |
| Retrograde fast axonal transport, squid | 2 | Individual dynein motors |
| Chromosome movement in anaphase of mitosis | 0.003–0.2 | Motors plus depolymerization |
| Endoplasmic reticulum sliding, Newt cell | 0.1 | Individual kinesin motors |
| Slow axonal transport, rat nerves | 0.002–0.1 net | Motors on microtubules |
| 1 (intermittent) | ||
| Microtubule Polymerization | ||
| Endoplasmic reticulum tip elongation, Newt cell | 0.1 | Microtubule polymerization |
| Actin-Myosin Motors | ||
| Cytoplasmic streaming, Nitella | 60 | Myosin motors on tracks |
| Cytoplasmic streaming, Physarum | 500 | Actin-myosin contraction |
| Actin Polymerization | ||
| Actin-propelled comet, Listeria | 0.5 | Actin polymerization |
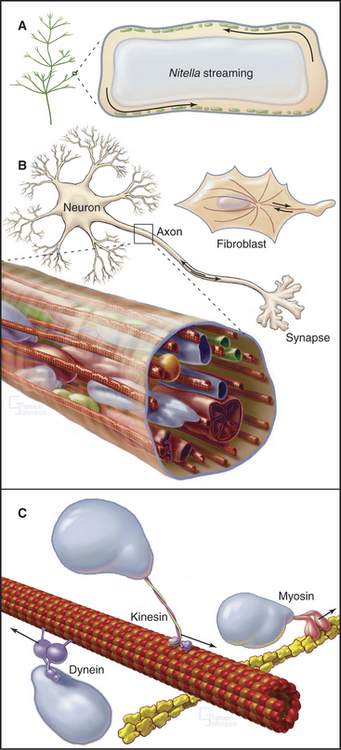
Two ancient mechanisms (Fig. 37-1) account for most intracellular movements in eukaryotes. Transport along microtubules by kinesin or dynein predominates in animal cells. Transport along actin filaments by myosin is more important in plants and fungi. Specialized isoforms of myosin, kinesin, and dynein are dedicated to particular movements. In most cases, transport involves single organelles, but vigorous organelle transport can even result in bulk movement of the cytoplasm. Alternatively, cytoplasmic contractions generated by myosin and actin filaments can produce cytoplasmic stream-ing, like squeezing toothpaste from a tube. Polymerization and depolymerization of microtubules and actin filaments also produce special types of intracellular movements.
Motor-driven movements on microtubules in animal cells generally receive the most attention, but this chapter takes a broad view across biology, highlighting all of the mechanisms for intracellular transport. Chapters on membrane traffic (see Chapters 20 to 21) and mitosis (see Chapter 44) cover more examples of intracellular movements.
Strategies to Identify Tracks and Motors
Historically, experiments with drugs that depolymerize or stabilize actin filaments or microtubules (see Boxes 33-1 and 34-1) provided the first clues about the cytoskeletal polymers that support various biological movements. Identifying the participating motors, if any, has been much more challenging, given a minimum five myosin genes, six kinesin genes, and one dynein gene in budding yeast and about ten times more motor protein genes in humans. A limited number of pharmacologic agents (Box 37-1) can implicate some motors, but the most definitive approach is to alter motor activity or abundance genetically or by RNAi-mediated depletion. Some motors are essential, so deletion mutations are lethal. Conditional mutations have allowed many definitive tests for the functions of these motors in yeast and, in a few cases, in more complex organisms. Other motors are not essential in the sense that cells have alternative strategies that can compensate if the protein is missing or inactive; nevertheless, the protein may have an important function. For example, dynein contributes to mitosis, but Drosophila tissue culture cells that are depleted of dynein can complete mitosis, though only after a long delay during which other motors take over.
Rapid Movements along Microtubules
Organelles in most cells are capable of moving at relatively high velocities, on the order of 1 mms (Table 37-1) along linear microtubule tracks. Thus, the organization of microtubules determines the patterns of these movements (Fig. 37-1; also see Fig. 34-2). Such movements are typically intermittent.
Microtubule-based motor proteins, kinesins and dyneins, power organelle movement along microtubules. Kinesins move organelles toward microtubule plus ends, which are located near the periphery of cells with microtubules radiating from centrosomes. Dyneins are responsible for organelle movements toward the minus ends of microtubules, located at the cell center. Intraflagellar transport of proteins in cilia and flagella (see Fig. 38-20) shares many features with the movements of organelles. Movements of organelles along microtubules have been reconstituted with purified dynein and kinesin. Final proof of the responsible motors usually depends on genetic tests.
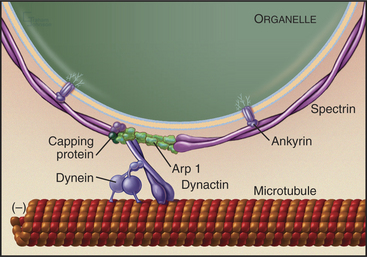
An assembly of proteins called the dynactin complex regulates the ability of dynein to transport membranes along microtubules (Fig. 37-2). This complex consists of a short filament of the actin-related protein Arp1 and seven other subunits, including heterodimeric capping protein. The 150-kD subunit (p150glued) binds to an intermediate chain of dynein. The Arp-1 filament interacts with spectrin associated with the membrane. Mutations in Drosophila p150glued cause developmental defects in the eye and brain. Some patients with inherited forms of motor neuron degeneration also have mutations of p150glued.
Fast Axonal Transport
High-contrast light microscopy of living axons reveals that most membrane-bound organelles move either toward (anterograde) or away from (retrograde) the end of the axon (Fig. 37-3) with some pauses and even occasional changes of direction. Retrograde movements (2.5 mm s−1 or 22 cm/day) are faster than anterograde movements (0.5 mm s−1 or 4 cm/day). At these rates, a round trip from a cell body in the spinal cord of a human to the foot and back takes only three weeks. This might seem slow, but if a 0.1-mm vesicle were the size of a small car, it would be moving anterograde at 50 miles per hour and retrograde at 250 miles per hour. In the axons of vertebrate neurons, mitochondria and autophagic vesicles move back and forth in both directions. Their net movement toward the nerve terminal or cell body depends on physiological conditions.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


