“The duck is the Trojan horse.”
—Dr. Robert G. Webster, influenza researcher, St. Jude Children’s Research Hospital
9.1 History of Influenza
Influenza is not a new disease. Historians described outbreaks of a highly contagious respiratory illness followed by pneumonia as early as 400 BC. The word influenza was derived from the Latin word influentia because it was believed diseases were caused by bad heavenly fluid, or “influence.” Influenza was used as a general term. For example, scarlet fever was referred to as influenza di febbre scarlattina, and catarrhal fever (a very bad head cold involving copious mucus production) was called influenza di catarro. The first word in the phrase became the most significant. The French term la grippe, or “the seizure,” became the most popular term during the early 1900s. In English, diseases correlated with most respiratory tract infections became known as influenza, later abbreviated to the modern flu.
Major influenza epidemics (localized outbreaks) and pandemics (worldwide epidemics) have been recorded for centuries. Pandemics occurred in 1918 (Spanish influenza; see VIRUS FILE 9-1 and Section 9.10), 1957 (Asian flu), 1968 (Hong Kong flu), 1977 (swine flu), and 2009 (H1N1 swine flu). Pandemics have occurred when influenza A virus mutates dramatically, resulting in high morbidity and mortality.
Today, millions of Americans contract influenza each year. CDC experts used to report that during the average flu season there were approximately 36,000 deaths nationwide and many more hospitalizations. Today, CDC experts state that the range of annual deaths is too wide to continue using a single number. Based on 31 seasons (1976–2007), the average is closer to 23,600, with a range of approximately 3,300–49,000 influenza deaths each year. The elderly, pregnant women, very young children, and otherwise immunocompromised individuals are at the highest risk of developing serious complications associated with influenza. This group accounts for more than 90% of influenza-related deaths. Influenza is the seventh leading cause of death in the United States. Many virologists think that another influenza pandemic that could kill millions of people is inevitable.
9.2 Epidemiology of Influenza
At any given time of the year, one or two strains of influenza A and B viruses dominate a particular region of the world. In temperate or colder regions, influenza illness usually occurs during the winter, but in tropical regions influenza occurs throughout the year. Influenza epidemics can become unmanageable at alarming speeds for several reasons:
The incubation period, also known as the time between infection and the first appearance of symptoms, is only 1–4 days.
Copious numbers of infectious virus particles are shed in droplets discharged by sneezing or coughing. One droplet can contain 100,000–1,000,000 virus particles. The infectious dose of influenza A virus is 1,000–2,000 infectious virions.
Many symptomatic individuals do not stay at home. Instead, they continue their normal activities, spreading the influenza virus to many contacts in the process.
Individuals lack immunity to the strain of influenza A or B virus circulating in the area at the time of the epidemic.
Schools are a primary source where influenza viruses are shared. Children bring the viruses home. Parents who contract influenza then disseminate the infection into the workplace. The best indicator distinguishing the scale of an epidemic is an escalation in absenteeism from schools and the workplace that is accompanied by an increase in hospital admissions and deaths, especially among the elderly. Epidemiologists measure the increase in mortality during an influenza epidemic by comparing it to the average number of deaths during comparable winters without an influenza epidemic. The number of “excess deaths” helps to determine epidemic years.
In the United States, Spanish influenza killed 675,000 Americans, with a national average of 4.39 deaths per 1,000 people. Death rates peaked during October 1918 (FIGURE 9-1A). Graphical analysis of the 1918 influenza pandemic shows a “W-shaped” curve in which mortalities peaked among the very young, the elderly, and those ages 20–40 years. The 1918 flu was unique in that it killed healthy adults between 20 and 40 years of age (FIGURE 9-1B) in addition to children and elderly, who are usually within the high-risk group. Typical influenza season deaths are represented graphically by “U-shaped” curves in which peak mortalities caused by influenza virus occur among the very young and the elderly (Figure 9-1b). The 1918 influenza pandemic decreased life expectancy in the United States by 11 years (FIGURE 9-1C).
In the United States, the 1957–1958 Asian flu caused 70,000 deaths and the 1968–1969 Hong Kong flu resulted in 34,000 deaths. The mortalities were caused by influenza type A viruses (see Section 9.4). Each flu season is unique, but it is estimated that about 10–20% of U.S. residents get the flu, and an average of 114,000 persons are hospitalized for flu-related complications during each flu season.
9.3 Clinical Features of Influenza
Influenza can be hard to diagnose because there are several other infectious diseases that resemble it. Accurate diagnosis is based on the patient’s history and physical examination in a clinical setting. Sometimes laboratory testing is performed before treatment is initiated.
Uncomplicated Infection
Patients, and even some doctors, have a tendency to label all respiratory ailments as “the flu.” In reality, influenza is a distinct clinical entity that begins after a 1- to 4-day incubation period. The illness starts with an unexpected onset of symptoms: headache; aching in the limbs and back; fever; malaise (a vague feeling of physical discomfort or uneasiness); a dry cough; sore throat, which can lead to the voice becoming husky and even lost; and myalgia (pain in muscles). Many patients can pinpoint the hour that they started feeling sick.
In general, most individuals feel weak and remain in bed. Their temperature will be high and continuous (100°–103°F [37.8°–39.4°C] for 3 days). Cough and weakness can persist for another week or more. Eyes are watery and burning and can even be painful upon movement. The nose may be runny or blocked with discharge. The patient’s eyes appear red, and his or her cheeks may be flushed. A chest x-ray will be normal unless a secondary bacterial infection has taken place. Usually the infection resolves within 7 days, although patients often complain of feeling listless for a few weeks after the infection.

FIGURE 9-1 (a) During the Spanish influenza pandemic of 1918, the month of October had the highest number of Spanish influenza deaths. (b) This graph compares the age distribution of influenza deaths in 1918 and 1911–1917. Influenza pandemics result in a “U-shaped” curve, with mortality peaks in the very young and elderly. The Spanish flu was unusual in that it also killed those in the prime of their lives, resulting in a characteristic “W-shaped” curve. (c) U.S. life expectancy dropped in 1918.
Many individuals think that the same virus causes the common cold and the flu. This is not the case. The common cold, or acute viral rhinitis, is a much milder disease that can be caused by one of over 100 different types of rhinoviruses and is not lethal. The differences between a common cold and influenza are listed in TABLE 9-1.
Complicated Infection
Complications are age dependent. Young children may develop croup (laryngitis with a hoarse, barking cough and difficulty breathing), secondary bacterial pneumonia, and middle ear infections. The elderly are particularly vulnerable to life-threatening pneumonia caused by secondary bacterial infections, usually due to Staphylococcus aureus, Streptococcus pneumoniae, or Haemophilus influenzae. Preexisting chronic conditions such as congestive heart failure can be exacerbated.
Besides the elderly, anyone suffering from chronic conditions affecting the pulmonary (lung), cardiac (heart), renal (kidney), hepatic (liver), or endocrine system is at risk. Modern medicine has kept many children alive who have immune deficiencies or diseases such as cystic fibrosis as well as saved patients of any age with organ transplants, cancer, or acquired immunodeficiency syndrome (AIDS). Medical advancements have increased the number of individuals at risk of death during an influenza epidemic.
Reye’s Syndrome
Besides secondary bacterial infections, a rare condition called Reye’s syndrome is sometimes associated with influenza (or other viral illnesses, such as chickenpox) in children. Studies have shown that using aspirin or salicylate-containing medications to treat viral illnesses increases the risk of developing Reye’s syndrome. It affects all organs of the body but is most harmful to the brain and the liver, causing an acute increase of pressure within the brain and massive accumulations of fat in the liver and other organs.
Symptoms of Reye’s syndrome include persistent or recurrent vomiting, listlessness, personality changes (i.e., irritability or combativeness), disorientation or confusion, delirium, convulsions, and loss of consciousness. There is no cure for Reye’s syndrome. Successful management depends on early diagnosis.
Anthrax, Influenza, SARS, or MERS?
After the September 11, 2011, terrorist attacks on the United States, clinicians were concerned about how to evaluate persons who may be at increased risk for inhalational anthrax but who have symptoms of an influenza-like illness. By November 2001, the CDC published a table listing the signs and symptoms of 10 cases of inhalation anthrax compared to laboratory-confirmed influenza. It is important to be able to distinguish among respiratory infections—especially during the flu season— so that they can be treated appropriately (TABLE 9-2).
Table 9-1 Common Cold or Influenza?
| Symptom | Common Cold | Influenza (Flu) |
|---|---|---|
| Fever | Rare in adults and older children, but can be as high as 102°F (39°C) in infants and small children | High (100°–104°F [38–40°C]), lasting 3–4 days. |
| Headache | Occasionally | Prominent, sudden onset |
| General muscle aches and pain | Slight/mild | Common, often severe |
| Sore throat | Sometimes | Common |
| Cough, chest discomfort | Mild hacking cough | Common, can become severe |
| Sneezing | Usual | Sometimes |
| Runny or stuffy nose | Common | Sometimes |
| Fatigue, weakness | Sometimes | Often extreme, lasting 2–3 weeks |
| Extreme exhaustion | Never | Sudden onset; can be severe |
| Complications | Sinus congestion or earache | Bronchitis, pneumonia |
| Prevention | Wash hands often; avoid close contact with individuals who are suffering from a cold | Annual vaccination, antiviral drug |
| Treatment | None, temporary relief of symptoms | Antiviral drug within 24–48 hours after onset of symptoms |
Table 9-2 Symptoms and Signs of Inhalational Anthrax vs. Laboratory-Confirmed Influenza, SARS, and MERS
| Symptoms/Signs | Inhalational Anthrax (n = 10)a | Laboratory-Confirmed Influenzaa | SARS, Globalb | MERS (n = 47)b |
|---|---|---|---|---|
| Fever or chills | 100% | 83–90% | 15–73% | 87% |
| Fatigue/malaise | 100% | 75–94% | 31–45% | 38% |
| Nonproductive cough | 90% | 84–93% | 29–75% | 47% |
| Shortness of breath or chest discomfort/pain | 80% | 6% | 40–42% | 72% |
| Headache | 50% | 84–91% | 20–56% | 13% |
| Sore throat | 20% | 64–84% | 13–25% | 21% |
| Runny nose | 10% | 79% | 2–24% | 4% |
| Nausea or vomiting | 80% | 12% | 20–35% | 21% |
| Abdominal pain or diarrhea | 30% | 22% | 20–25% | 26% |
| Myalgia (muscle pain) | 50% | 67–94% | 45–61% | 32% |
aThe CDC reported the signs and symptoms of 10 cases of inhalational anthrax that occurred after the tragic events of September 11, 2001, and compared them to laboratory-confirmed influenza (data from MMWR, 2001, 50[44]:984–986). The inhalational anthrax cases began appearing just before influenza season, raising diagnostic concerns for healthcare providers. Many persons who were exposed to inhalational anthrax in the bioterrorism-related anthrax attacks completed a 60-day course of antibiotic prophylaxis. bData comparing SARS and MERS from Assiri, A., et al. 2013. “Epidemiological, demographic, and clinical characteristics of 47 cases of Middle East respiratory syndrome coronavirus disease from Saudi Arabia: A descriptive study.” Lancet Infect Dis 13:752–761. | ||||
In 2013, a study was published to distinguish severe acute respiratory syndrome (SARS) from Middle East respiratory syndrome (MERS), a new human respiratory illness first reported in Saudi Arabia in 2012. SARS originated in southern China in 2002, spread to Hong Kong, and then rapidly spread globally, resulting in 8,422 cases and 916 deaths (11% mortality). Like SARS, MERS is caused by a novel coronavirus. It shares similar clinical features with SARS. Most patients with serious symptoms, such as shortness of breath, have either preexisting conditions or immunosuppression (Table 9-2).
9.4 Classification of Influenza Viruses
True influenza is caused by any of several genera of the Orthomyxoviridae family of viruses. Orthos is the Greek term meaning “standard” or “correct,” and myxa means “mucus.” Orthomyxoviridae contains three influenza genera categorized as influenza type A, influenza type B, and influenza type C viruses. All three types of influenza viruses can cause similar symptoms in humans. Infection with one type of influenza, however, does not confer immunity to the other types. TABLE 9-3 compares the epidemiological, clinical, genomic, and structural differences among the three virus types. All three share certain structural features, including segmented genomes composed of single-stranded RNA (ssRNA) of negative polarity and a host-derived envelope containing glyco-proteins critical for virus entry and exit of host cells.
Table 9-3 Differences Among Influenza Types A, B, and C Viruses
| Feature | Influenza A Virus | Influenza B Virus | Influenza C Virus |
|---|---|---|---|
| Host range | Humans, pigs, horses, birds, marine mammals | Humans only | Humans and pigs |
| Epidemiology | Antigenic shift and antigenic drift | Antigenic drift only | Antigenic drift only |
| Clinical features | May cause pandemics with significant mortalities in affected young people | Severe disease, generally confined to elderly or high-risk; pandemics not seen | Mild disease, common in children, without seasonality |
| Genome | 8 gene segments | 8 gene segments | 7 gene segments |
| Structure | 10 viral proteins; M2 unique | 11 viral proteins; NB unique | 9 viral proteins; HEF unique |
9.5 Laboratory Diagnosis of Influenza
Most physicians use multiple signs and symptoms to diagnose influenza, backed up by the knowledge that an outbreak is in progress. Influenza viruses, common cold viruses, and bacteria can cause similar upper respiratory tract symptoms. The intensity, severity, and frequency of the symptoms vary. FIGURE 9-2 illustrates the natural course of influenza infection. When a tentative diagnosis of influenza is made, laboratory testing may be an appropriate confirmatory step.
Office-Based Rapid Viral Tests
The U.S. Food and Drug Administration (FDA) has approved at least 19 commercially available, office-based techniques for detecting influenza type A and/or B. All of the rapid tests can be completed within 10–20 minutes. Rapid influenza tests are cheaper than virus isolation: ~$15 per test (kits contain 25 tests/box). The rapid influenza tests are performed using nasal swab specimens and are either enzyme-linked immunosorbent assays (ElIsAs) or immunofluorescent assays. Of the 19 FDA-approved rapid diagnostic tests, 7 require an analyzer reader device. If the chosen test is positive, antiviral therapy may be considered if symptoms have been present for less than 48 hours. A negative test result may support the decision to withhold antiviral drug therapy. Although rapid tests are relatively easy to perform, the CDC estimates that up to 30% of samples may produce negative results yet be positive by viral culture methods. Rapid tests may also produce false-positive results.
Real-Time PCR Diagnostics
Pig farmers now have access to a rapid molecular diagnostic kit that screens nasal swab specimens from pigs for swine influenza virus. VetMAX-Gold SIV Subtyping Kit is the only diagnostic kit licensed by the United States Department of Agriculture (USDA). The detection kit uses real-time polymerase chain reaction (real-time PCR) technology that is rapid and cost-effective. It can be used in combination with other kits that can distinguish influenza virus subtypes.

FIGURE 9-2 The natural course of influenza in an otherwise healthy 28-year-old male patient. Many symptoms overlap throughout the duration of the respiratory illness. Influenza viruses are shed during the first 5 days of infection.

FIGURE 9-3 Animal and Plant Health Inspection Service (APHIS) veterinary technician completes the Swine Health Protection Program premises inspection form.
Swine influenza is highly contagious; it causes fever, lethargy, and weight loss in pigs, resulting in economic losses for producers (FIGURE 9-3). It is virtually impossible to maintain hogs that are free of influenza A virus infections. No specific treatment is available for swine influenza. Farmers may administer antibiotics to sick sows to control or prevent secondary bacterial infections.
A real-time PCR influenza virus A and B assay that can be completed in 20 minutes was approved in 2015 for use in pharmacy clinics, doctor’s offices, and other health clinics. The entire testing process is automated, from sample prep to the real-time PCR reactions that are performed by a small benchtop instrument called the cobas Liat PCR System (Roche Diagnostics).
Conventional Virus Isolation Detection in Cell Cultures
Propagating or isolating viruses in cell cultures is the most accurate method for identifying specific viral strains. Virus can be recovered from nasal swab specimens, sputum samples, nasal washes, or combined nose-and-throat swab specimens. The collected clinical specimens are placed in special transport containers, labeled, and transported properly to the viral diagnostics lab as soon as possible.
Antibiotics may be added to the clinical specimens to inactivate contaminating bacteria prior to inoculating rhesus or cynomolgus monkey kidney cells or Madin-Darby canine kidney (MDCK) cell cultures. Influenza viruses are detected based on microscopic observation of cytopathic effects (CPEs) in the cell cultures or other assays. Results are not available for 2–10 days. Viral culture is also costly (about $75) and requires special laboratory procedures as well as skilled technicians to carry out these procedures. In sophisticated laboratories, nucleic acid techniques such as PCR; viral antigen detection through serology methods; or visualization of virions using electron microscopy, cytology, or histology may be used to diagnose viral illness.
Serology Testing
Sometimes in clinical practice blood is drawn and serum from the patient is used for diagnosis to detect antibodies against influenza virus. Blood is drawn again after the patient has recovered, usually 10–20 days later, and convalescent serum is screened for viral antibodies. The serum samples are analyzed for an increase in antibody titer. A fourfold or greater increase in antibody titer is considered diagnostic of viral infection. The diagnostic information is collected too late, and therefore does not affect clinical decisions; however, it does confirm evidence of the viral infection.
Microarray “Flu Chips”
Over 20 different types of nucleic acid tests have been approved by the FDA for the detection of influenza and respiratory viruses. Some of the molecular tests use “flu chips,” or microarrays, as a way to quickly identify a variety of respiratory pathogens within an hour or less. The microarrays screen for influenza types A, B, and C viruses; avian influenza viruses; severe acute respiratory syndrome coronavirus (SARS-CoV); respiratory syncytial virus (RSV); and other airborne bioterrorist pathogens, such as Bacillus anthracis. Several teams are developing diagnostic microarrays that address surveillance of influenza viruses at U.S. pig markets and the needs of developing nations by creating inexpensive and field-portable test kits for influenza A virus respiratory tract infections.
When a U.S. public health emergency response to H1N1 influenza A was declared in December 2009, the FDA approved the temporary use of resequencing influenza A microarray detection panels to diagnose infections caused by the 2009 influenza A virus (H1N1) through April 26, 2010.
9.6 Cellular Pathogenesis
Influenza is spread person to person through droplets containing influenza viruses. Large numbers of influenza viruses are present in respiratory secretions of infected persons. Influenza viruses are dispersed in aerosol form by sneezing, coughing, and talking. Infection occurs through the inhalation of aerosolized infectious influenza particles. After influenza viruses enter the respiratory tract, viruses can attach to and penetrate ciliated columnar epithelial host cells lining the sinuses and airways (FIGURE 9-4A).

FIGURE 9-4 (a) Cilia are attached to columnar epithelial cells. Cilia sweep mucus produced in goblet cells as well as mucus coming from deeper glands within the lungs and the particulate matter trapped in the mucus. The presence of cilia aids the removal of mucus materials from lower parts of the airway to the throat for disposal. Influenza A viruses infect epithelial cells lining the respiratory tract. (b) Influenza A and other respiratory viruses enter and exit host cells that line the nasopharynx. Influenza A viruses also enter host cells that line the tracheobronchial tree.
The primary site of infection occurs at the tracheo-bronchial tree, but the nasopharynx is also involved (FIGURE 9-4B). After influenza virus attaches to the outside of a host ciliated epithelial cell, it is adsorbed and viral replication begins. The ciliated epithelial cell is destroyed, releasing progeny influenza viruses that spread to nearby cells. As the number of cilia are reduced, there are fewer to clean the lungs, and more mucus stays in the airways, clogging them and making the individual cough. Hence, the cleaning system in the lungs does not work well. The loss of epithelial cells and destruction of cilia may contribute to secondary bacterial invasion, manifested as sinusitis, otitis media (inflammation of the ear causing an earache), and pneumonia (infection of the alveoli and surrounding lung). Infectious influenza viruses can be isolated for 1–7 days, with the peak of the released virions occurring on days 4 and 5 after infection (FIGURE 9-5).
9.7 Immunity
Infection with influenza A virus results in the development of antibodies against the hemagglutinin (H) and neuraminidase (n) envelope glycoproteins present on the surface of the virus (see Section 9.8). Anti-H antibodies neutralize influenza A virus, preventing attachment/viral infectivity. Antibodies against N do not neutralize influenza A virus, but they do reduce release of virions from infected host cells. If an individual suffered from influenza within the past couple of years caused by a closely related strain of influenza A virus H subtype, anti-H IgA class antibodies may intercept and neutralize infectious virions. IgG antibodies may play a secondary role in neutralizing influenza A virus, providing protection in the lung.
If preexisting antibodies are inadequate, the body must mount an antibody response against influenza A virus. Besides an antibody response, other players of the immune system play a role in recovery, such as activated macrophages; TC lymphocytes; and certain cytokines, such as interferon.

FIGURE 9-5 Time course of infections caused by influenza A viruses. Flulike symptoms occur early. Later, pneumonia may result from a secondary bacterial infection in certain individuals.
A cytokine storm, or systemic inflammatory response syndrome (SIRS), was suggested as an explanation for the devastating physiological effects of the 1918 strain of influenza A virus on the respiratory system of its victims. From a clinical perspective, a cytokine storm is defined as an immune system that is overreacting toward the pathogen. Cytokine-signaling immune cells such as macrophages and T lymphocytes travel to the site of infection, especially the lungs, causing damage that ultimately results in failure of multiple organs (VIRUS FILE 9-2). SIRS is rare. Patients who suffer from this type of immune response usually die. The 1918 influenza A H1N1 strain was re-created using published sequences of the viral genome and reverse genetics (see Section 9.10 and Virus File 9-3). The re-created virus was highly virulent in macaques and mice. Scientists determined that the following chemokines or cytokines were present at elevated levels in the bronchi of macaques infected with the 1918 influenza A virus: interleukin 6 (IL-6), interleukin 8 (IL-8), monocyte chemotactic protein 1 (MCP-1 or CCL2), and CCL5 (RANTES). The Asian avian influenza A (H5N1) virus that crossed the species barrier to infect humans also causes SIRS and has a 50% mortality rate in humans.
9.8 Influenza A Virus Replication
Influenza a Virus Structure
Influenza A virus particles are 80–120 nm in diameter. Influenza A virions may be filamentous in a form of helical symmetry immediately after isolation from cell cultures in the laboratory (FIGURE 9-6A). After several passages in cell cultures, the morphology of influenza A virions becomes spherical (FIGURE 9-6B). Transmission electron micrographs reveal influenza A virus to be enveloped. Its surface is covered with approximately 500 glycoprotein spikes projecting outside of the host-derived viral envelope. Approximately 80% of the rod-shaped glycoprotein spikes are hemagglutinin (H or HA), and 20% are mushroom-shaped neuraminidase (N or NA). The H is a homotrimeric integral membrane glycoprotein, whereas N is a homotetrameric integral membrane glycoprotein (FIGURE 9-6C). Matrix 1 (M1) proteins form a layer beneath the viral membrane, surrounding the eight segments of the viral genome. A few copies of nuclear export proteins (NS2) are also found inside of the virion.
In addition to the integral N and H proteins found on the surface of influenza A virus, there are a few M2 ion channel integral membrane proteins. M2 is not present in influenza B virions. M2 ion channel proteins are inserted into the lipid bilayer membrane that surrounds the nucleocapsid of the virus (nucleoproteins + nucleic acids). M1 is the most abundant virion protein. The virus particle contains eight segments of ssRNA of negative polarity (–ssRNA), ranging from 934 to 2,341 nucleotides in length (TABLE 9-4). The –ssRNA segments are coated with nucleoproteins (NP). The coding assignments for each segment are depicted in the schematic diagram of the influenza A virus in FIGURE 9-7. Each segment contains an RNA-dependent RNA polymerase or transcriptase complex that consists of three additional viral polymerase proteins: PB1, PB2, and PA (Figure 9-6c). Nonstructural proteins (NS1) are only found in infected cells.
Virus Nomenclature
The standard naming for influenza A viruses is as follows:
Influenza type/
Species isolated from/ (unless human)
Place of isolation/
Strain designation/
Year of isolation/
H#N# subtypes
Consider the following example: initial influenza A viruses isolated from a clinical specimen collected from a patient suffering from influenza in Hong Kong during a 1968 influenza outbreak would be named A/Hong Kong/1/68(H3N2). If the strain is a human strain, the host origin is not included in the name. However, if the influenza A virus isolate was isolated from another host, such as ducks, the host name would be A/duck/Hong Kong/15/68(H1N1). Influenza virus type A is the most frequent cause of influenza and is the focus of this chapter.

FIGURE 9-6 Transmission electron micrographs of influenza A viruses. Note the glycoprotein spikes protruding from the surface of the viral particle. (a) When influenza A viruses are isolated directly from patients, the virions are filament shaped, as shown in the transmission electron micrograph. (b) After several passages in the laboratory, the virion morphology of influenza A viruses becomes spherical shaped. (c) Schematic diagram of influenza A virus structure. H and N spikes protrude from the enveloped virion. H is a trimer of identical subunits. N is a tetramer of identical subunits. The ratio of H to N varies, but it ranges from 4:1 to 5:1. The viral envelope also contains a few M2 ion channel transmembrane proteins. The virion contains a genome consisting of eight segments of –ssRNA. A transcriptase complex composed of PB1, PB2, and PA viral proteins is bound to the ends of each –ssRNA segment. The genome is protected by NP and M1 matrix proteins. NS2 is also packaged into the virion.
There are 18 antigenically different types of Hs (H1–H18 subtypes) and 11 distinct Ns (N1–N11 sub-types). All subtypes differ by more than 30% amino acid sequence homology. All H and N subtypes infect waterfowl (e.g., ducks). Six subtypes of H (H1, H2, H3, H5, H7, and H9) and two subtypes of N (N1 and N2) have been isolated from humans (TABLE 9-5). All but the H5, H7, and H9 subtypes spread efficiently by human-to-human contact. All influenza A virus subtypes circulate in wild waterfowl (mainly ducks, geese, and swans) and shore-birds (mainly gulls, terns, and waders), which are considered the natural reservoir of influenza A viruses, with the exception of H17, H18, N10, and N11. Influenza A viruses H17N10 and H18N11 subtypes were recently identified in rectal swabs of yellow-shouldered and flat-faced fruit-eating bats, raising the possibility that birds are not the exclusive influenza A virus reservoir.
Besides humans, birds, and bats, other H and N sub-types are found in wild or domesticated animals such as seals, whales, camels, mink, palm civets, dogs, tigers, leopards, stone martens, cats, and horses (Table 9-5; see also Section 9.9 and Figure 9-14).
Cross-reactivity among the different subtypes does not occur. In other words, if you are immune to influenza A virus (H1N1), you will not have any immunity against an H2N2 subtype. The antigenic subtype differences are critical for vaccine preparation (see Section 9.13).
Table 9-4 Influenza A Virus Gene-Coding Assignments
| RNA Segment | Length (Bases) | Gene Product | Function |
|---|---|---|---|
| 1 | 2,341 | PB1 757 | Catalyzes nucleotide addition (part of transcriptase complex) |
| 2 | 2,233 | PA 716 | Part of transcriptase complex |
| 3 | 2,341 | PB2 759 | Recognizes cap of host mRNAs, endonuclease (part of transcriptase complex) |
| 4 | 2,073 | HA 566 | Hemagglutinin Major surface glycoprotein Sialic acid binding Fusion at low pH |
| 5 | 1,565 | NP 498 | Nucleocapsid protein, interacts with RNA segments Involved in switching of mRNA to template RNA synthesis and virion RNA synthesis |
| 6 | 1,413 | NA 454 | Surface glycoprotein Neuraminidase (cleaves sialic acid) |
| 7 | 1,027 spliced | M1 252 M2 97 | M1 matrix protein inside of lipid envelope of virion M2 has H+ ion channel activity |
| 8 | 934 spliced | NS1 230 NS2 121 | NS1, nonstructural protein (not detected in virions), regulates viral RNA and protein synthesis and RNA splicing; also an IFN antagonist NS2 plays a role in nuclear export of ribonucleoproteins (RNPs) |
Genome Organization
Influenza A virus contains eight segments of ssRNA of negative polarity (see Table 9-4 for coding assignments). The complete nucleotide sequences of influenza A virus PR/8/34 and many RNA segments of other subtypes were completed in 1982. Segments 1–6 code for only one viral protein; segments 7 and 8 code for two.
The Virus Replication Cycle
Virus Adsorption and Entry
The influenza A virus H attaches to sialic acid residues that are present on glycoproteins or glycolipids of the ciliated columnar epithelial cells lining the sinuses and airways. Virions enter the cell within an endosomal vesicle by endocytosis (FIGURE 9-8A). Inside of the endosome, the influenza A virus encounters a relatively low pH (the pH drops from 7 to 5). The lower pH change triggers fusion between the viral lipid membrane and the host cell endosomal membrane (FIGURE 9-8B). The H protein mediates the low pH–dependent fusion event. The membrane fusion step is a vital step in the replication cycle of influenza A viruses. An irreversible conformational change of the H protein is triggered by the low pH of the host cell endosome. The H protein is composed of two polypeptides, designated H1 and H2. The full-length H protein is referred to as H0. The H1 and H2 polypep-tides are linked by a disulfide (S–S) bond between two cysteine amino acids. The H1 portion of H0 contains sialic acid–binding sites for attachment to the host cell. The H2 portion of H0 forms a membrane-spanning anchor or a fusion peptide, which is directly involved in the fusion mechanism (Figure 9-8b). The H2 peptide interacts with the host membrane after the conformational change is induced in H0 by low pH.

FIGURE 9-7 The genome of influenza A virus consists of 8 RNA segments, coding for 10 influenza protein products. Gene segments 7 and 8 code for more than one protein on overlapping reading frames and the viral RNAs undergo a splicing event.
Table 9-5 Hemagglutinin (H) and Neuraminidase (N) Subtypes of Influenza A Viruses Isolated from Humans, Domesticated Animals, Birds, and Bats
| Subtype | Human | Swine | Horse | Bird | Bat |
|---|---|---|---|---|---|
| Hemagglutinin | |||||
| H1 | √ | √ | √ | ||
| H2 | √ | √ | √ | ||
| H3 | √ | √ | √ | √ | |
| H4 | √ | ||||
| H5 | √ | √ | |||
| H6 | √ | √ | |||
| H7 | √ | √ | √ | ||
| H8 | √ | ||||
| H9 | √ | √ | |||
| H10 | √ | ||||
| H11 | √ | ||||
| H12 | √ | ||||
| H13 | √ | ||||
| H14 | √ | ||||
| H15 | √ | ||||
| H16 | √ | ||||
| H17 | √ | ||||
| H18 | √ | ||||
| Neuraminidase | |||||
| N1 | √ | √ | √ | ||
| N2 | √ | √ | |||
| N3 | √ | √ | |||
| N4 | √ | ||||
| N5 | √ | ||||
| N6 | √ | ||||
| N7 | √ | √ | |||
| N8 | √ | √ | |||
| N9 | √ | √ | |||
| N10 | √ | ||||
| N11 | √ | ||||
See reviews on influenza A virus subtypes in humans, animals, marine mammals, domesticated animals, wild and domesticated birds, reptiles, amphibians, and bats: Reperant, L. A., et al. 2009. “Avian influenza viruses in mammals.” Rev Sci Tech 28:137–159; Short, K. R., et al. 2015. “One health, multiple challenges: The inter-species transmission of influenza A virus.” One Health 1:1–13. Information from Levine, A. J. Viruses. W. H. Freeman and Company, 1992. | |||||

FIGURE 9-8 Influenza A virus attachment and entry. Influenza A viruses attach to sialic acid receptors present on the surface of ciliated epithelial cells lining the sinuses and airways. (b) Conformational changes within the influenza A virus H protein trigger fusion of the virion membrane with the cellular membrane. (c) The sequence of the cleavage site determines which proteases (intracellular or extracellular) cleave H0. Influenza A viruses are not infectious unless H0 is cleaved by proteases into H1 and H2.
Influenza A virus is not infectious unless the H protein is cleaved by cellular proteases. If the H protein contains a furin cleavage site, it undergoes proteolytic cleavage by a cellular protease inside of the trans-Golgi network. If it does not contain a furin cleavage site, the H0 precursor protein is cleaved into H1 and H2 by cellular proteases outside of the host cell. This limits their spread in hosts to tissues where the appropriate proteases are encountered. FIGURE 9-8C is a schematic of the cleavage site within the protein.
A general rule has been observed. The H proteins of pathogenic influenza A viruses are cleaved intracellularly by proteases and have the capacity to infect various cell types and cause systemic infections. The H proteins of influenza A virus that are cleaved by extracellular trypsin-like proteases are generally apathogenic, limiting their spread in hosts to tissues that contain the appropriate proteases. During some influenza A virus infections, the proteases of coinfecting bacteria may play a role in influenza A virus activation and cause an increase in the severity of the respiratory illness.
Uncoating Step
uncoating takes place in the host cell endosome. After the viral membrane fuses with the endosomal membrane of the host cell, the viral M2 transmembrane protein forms a transmembrane H+ ion channel or small pore in the viral envelope. The M2 ion channel protein allows H+ ions to penetrate the virion, weakening the interaction of the viral M1 matrix protein from the viral RNA, NP, and transcriptase or RNA-dependent RNA polymerase complex (RNP). The RNPs are released into the cytoplasm and are exported to the nucleus. The antiviral drugs amantadine (sold as Symmetrel) and rimantidine (sold as Flumadine) block the M2 ion channel function, resulting in the incomplete release of the viral RNPs, ultimately interfering with uncoating (FIGURE 9-9 and Section 9.12, Antivirals for Influenza Treatment).

FIGURE 9-9 Fusion and uncoating of influenza A viruses. M2 facilitates the movement of protons from the endosome into the inside of the virion. The ribonucleoproteins (RNPs) are liberated into the cytoplasm and transported to the host cell nucleus. Two drugs, amantadine and rimantidine, block M2 ion channel activity, preventing viral uncoating.
mRNA Synthesis and Replication of Virion RNA
Viral genome replication of influenza A virus occurs in the nucleus of the infected cells. This is an unusual feature for an RNA virus because the genome of most RNA viruses is replicated by RNA-dependent RNA polymerases in the cytoplasm of the infected cell. Following attachment, entry, and uncoating, RNPs are transported to the nucleus and viral mRNA synthesis begins. During synthesis of mRNA, influenza A virus engages in cap snatching. The viral PB2 protein binds to the cap structures of host mRNAs located in the nucleus of the host cell. The cap, together with 10–13 nucleotides from the 5′ end of the host mRNA, is removed, and PB1 and PA of the RNP complex use the short string of nucleotides as a primer to initiate transcription of viral RNA (vRNA) by extending the primer (FIGURE 9-10A). For this reason, influenza A viruses can only replicate in physiologically active cells containing a functional DNA-dependent RNA polymerase (RNA polymerase II) even though this enzyme does not transcribe the virus genome.
Transcription of the viral mRNA is terminated near the end of the genome segment at a point on the template vRNA that contains a run of uracil residues 17–22 residues from its 5′ end. These are copied into A residues in the mRNA, generating a poly(A) tail before the mRNA dissociates from the vRNA template. Synthesis of the antigenome positive-sense RNA does not use a primer and generates a complete complementary copy of the template RNA, termed vcRNA (FIGURE 9-10B). Two types of viral RNAs are generated: a positive-sense viral mRNA and a positive-sense viral antigenome RNA. TABLE 9-6 distinguishes between these two types of viral RNAs.
The Caps of Host mRNAs Are Snatched by Influenza A Virus
Interestingly, the viral polymerase complex selectively “snatches” caps from cellular pre-mRNAs but not viral mRNAs. The selectivity is essential because if the 5′ ends of the viral mRNAs were removed and utilized as primers, overall synthesis and translation of viral mRNAs would not occur. How has this selectivity evolved? While studying the splicing mechanism of RNA segments 7 and 8 of influenza A virus, researchers discovered that the viral polymerase complex binds to the specific sequence 5′-AGCAAAAAGCAGG-3′ that is found in all mRNAs complementary to nucleotides 1–12 of the 3′ end of each vRNA segment. It protects the 5′ ends of all viral mRNAs but not cellular pre-mRNAs from cleavage by the polymerase complex (Figure 9-9b).
Table 9-6 The Differences Between the Influenza A Virus mRNA and Antigenome
| + Viral mRNA (+ vmRNA) | + Antigenome RNA (+vcRNA) |
|---|---|
| Shorter than template genome segment | Exact copy of the genome segment |
| Contains 3′ poly(A) tail | No 3′ poly(A) tail |
| Contains 5′ cap | No 5′ cap |
| Synthesis is insensitive to inhibitors of protein synthesis | Synthesis requires continuous viral protein synthesis |
Influenza Exploits Host Nuclear Splicing Machinery
After the “snatched” host mRNA cap and associated nucleotides are used as a primer to transcribe each of the eight negative-sense RNA genome segments, six newly transcribed viral mRNAs (segments 1–6) are exported to the cytoplasm and translated immediately into viral proteins by the host translational machinery. Two of the primary viral transcripts (segments 7 and 8) are each spliced through the exploitation of the host splice machinery into at least two different viral mRNAs. FIGURE 9-11 is a schematic of the different viral mRNAs generated from influenza A virus segments 7 and 8. In both cases, the RNA splicing events remove a large portion of the first open reading frame (ORF), leaving the AUG initiation codon in place. The first alternative ORF is fused to the initiation codons of the M1 and NS1 ORFs. The newly generated ORFs direct the synthesis of novel proteins M2, NS2, and M3. All of these viral mRNAs code for nonstructural proteins. M3, however, has not been detected in infected cells, and its role in the infectious replication cycle is not known.
NS1 Prevents Nuclear Export of Cellular Pre-mRNAs, Facilitating Cap Snatching
Two cellular proteins—cleavage and polyadenylation specificity factor (CPSF) and poly(A)-binding protein II (PABII)—are responsible for processing the 3′ end of cellular mRNAs. Influenza A virus NS1 protein, the most abundant viral protein in infected cells, interacts with CPSF and PABII, thereby inhibiting their function. As a result, cellular pre-mRNAs are not cleaved and contain very short 3′ poly(A) tails that accumulate within the nucleus of the infected cell.
These newly synthesized cellular pre-mRNAs are trapped in the nucleus of the infected cell and are almost completely degraded. Retaining cellular pre-mRNAs in the nucleus facilitates the removal of the pre-mRNA 5′ cap by the influenza A virus PB2 protein. The host mRNAs synthesized after infection do not survive very long in the cell, and little or no cellular proteins will be synthesized in cells infected with influenza A virus.
Even though cellular pre-mRNAs are not exported to the nucleus, viral mRNAs can be exported and are not affected by this because the influenza A viral transcriptase complex adds the poly(A) tails onto the 3′ ends of viral mRNAs. The cellular CPSF and PABII proteins are not required for processing the 3′ ends of viral mRNAs (FIGURE 9-12).

FIGURE 9-10 (a) Cap-snatching mechanism of influenza A viruses. Three influenza proteins are involved in stealing the cap of host mRNAs: PB2, PB1, and PA. The cap, along with 10–13 nucleotides from the 5′ end of cellular mRNA, is removed from the host mRNA by PB1. RNA from the 5′ end of the cleaved cellular mRNA is used as a primer to initiate transcription of viral RNAs by the viral RNA–dependent RNA polymerase or transcriptase complex. (b) Schematic diagram distinguishing the differences between influenza A virion (–) genome RNA; viral (+) mRNA containing the snatched cellular mRNA cap structure; and (+) viral complementary RNA or template RNA that is a complete copy of the genome RNA segment.

FIGURE 9-11 (a) Splicing of RNA segments 7 and 8 of influenza A viruses. Shaded areas represent coding regions in either the 0, +1, or +2 open reading frames (ORFs). Thin lines represent untranslated regions. Introns are shown by the V-shaped lines. The first ORF codes for unspliced M1 and NS1 proteins. No evidence has been obtained that the M3 mRNA is detected in infected cells. Note that the M2 and NS2 proteins share common amino terminal sequences to the M1 or NS1 protein products, respectively.
Translational Control Mechanisms
Influenza A virus suppresses the interferon response to viral infection in host cells. During influenza A virus replication, double-stranded RNA (dsRNA) intermediates are formed. In a typical cellular interferon response, dsRNA molecules would activate the cellular kinase PKR, but influenza A virus counteracts PKR with a two-pronged attack:
The NS1 protein binds to the dsRNA intermediate molecules of influenza A virus produced during replication, sequestering them, and thereby blocking the activation of PKR.
Influenza A virus activates a 58-kDa cellular protein that interacts with PKR directly, inhibiting PKR in the infected cells.
Virion Maturation, Assembly, and Release
After the capped viral mRNAs are exported to the cytoplasm of the host cell, they are translated by ribosomes. The H, N, and M2 glycoproteins are synthesized on membrane-bound ribosomes. Following synthesis, these three membrane proteins enter the endoplasmic reticulum, where they are folded and glycosylated and subsequently transported to the Golgi network and the cell surface, where assembly takes place. At least one copy of each genome segment must be packaged into a new virion for it to be infectious.
The influenza A virions are assembled at the membrane surface of the host cell and released by a budding process. H anchors influenza A virus to the cell by binding to sialic acid receptors on the cell surface. The neuraminidase activity of N destroys sialic acid receptors present on the host cells during virus release or exit, preventing newly assembled viruses from aggregating or clumping together on the cell surface. Ultimately, N facilitates the release of viral progeny. It is also believed that N may aid influenza A viruses at the beginning of the replication cycle by aiding transport through the mucin layer in the respiratory tract so that they can attach to host epithelial cells. Zanamivir (Relenza), oseltamivir phosphate (Tamiflu), and peramivir (Rapivab) are antiviral compounds that inhibit the function of N (Figure 9-12 and Section 9.12, Antivirals for Influenza Treatment).
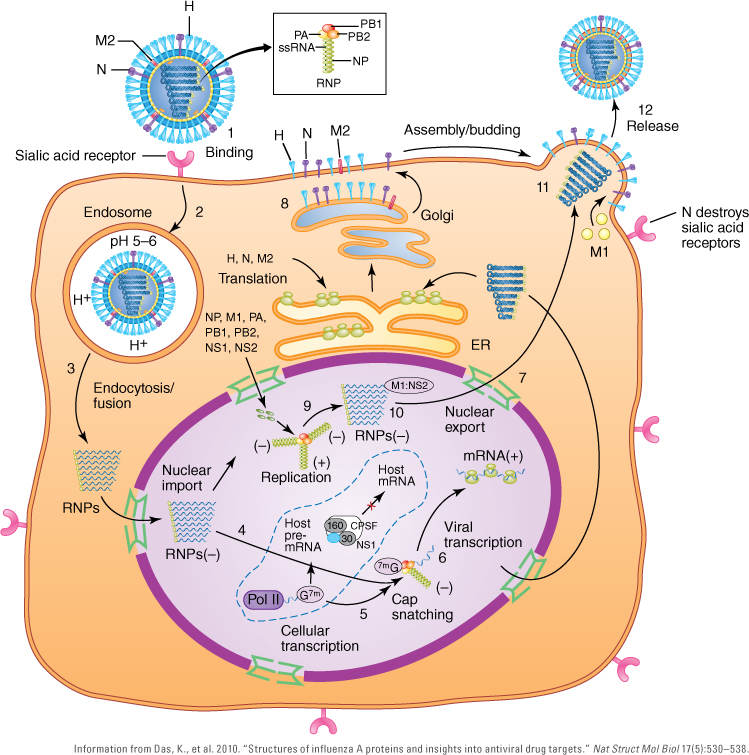
FIGURE 9-12 Replication cycle of influenza A virus. Step 1: The influenza A virus hemagglutinin (H) attaches to sialic acid host cell receptors. Step 2: The virus enters the host cell by endocytosis. Step 3: The M2 ion channel of the virus allows H+ ions to penetrate the virus. The low pH of the endosome triggers a conformational change in H, resulting in fusion of the viral and host endosomal membranes. Step 4: As the H+ ions enter through the M2 ion channel, the encapsidated genomic RNA or viral ribonucleoprotein complexes (vRnPs) are dissociated from M1 matrix proteins and released into the cytoplasm where they travel through the nuclear pores into the nucleus. Step 5: In the nucleus, the viral polymerase initiates its mRNA synthesis by cleaving, or “snatching,” 5′ caps from host pre-mRNAs. Step 6: PB2 binds to the 5′ cap of host pre-mRNAs, and PA endonuclease cleaves the host pre-mRNA 10–13 nucleotides downstream of the 5′ cap of the host pre-mRNA. Step 7: Replication/transcription of the viral genome is initiated. Step 8: Viral mRNAs are transported into the cytoplasm for translation into proteins. Step 9: Surface proteins—H, N, and M2—are processed in the endoplasmic reticulum (ER), glycosylated in the Golgi apparatus, and transported to the host cell plasma membrane. Step 10: NS1 suppresses host mRNA synthesis by inhibiting 3′-end processing of host premRNAs. Consequently, host mRNAs, including mRNAs that code for interferon, are blocked from synthesis, but viral mRNAs are unaffected by NS1 activity. Besides synthesizing viral mRNAs, the viral transcriptase complex is involved in the replication of genomic (–) vRNAs through a replicative intermediate, (+) vcRNA. Step 11: The final (–) vRNAs containing the transcriptase complex are encapsidated with M1, and NS2 is involved in their export through the nuclear pores to the site of virion assembly near the host cell plasma membrane. Newly assembled viruses exit by budding through the plasma membrane. Step 12: Newly assembled influenza A viruses are released as the neuraminidase (N) destroys sialic acid receptors on the surface of the host cell.
9.9 Genetic Variation in Influenza Viruses
mutations are more frequent in RNA viruses because their RNA-dependent RNA polymerases lack proofreading ability. RNA polymerases are at least 1,000–10,000 times more prone to error than human or viral DNA polymerases that are capable of proofreading during genomic replication. DNA polymerase proofreading is 3′ → 5′ exonuclease activity that removes misincorpo-rated nucleotides during DNA replication.
Mutations are common during influenza A virus genomic replication. The mutation rate of influenza A virus is approximately 1.5 × 10–5 mutations per nucleotide per replication cycle. It results in about two or three amino acid changes in the H protein each year. This rate is high compared to the mutation rate of mammalian genomes in which DNA polymerase has proofreading ability, resulting in 10–8 to 10–11 mutations per incorporated nucleotide per replication cycle.
Often there are “hot spots” for mutation in the viral genome. Influenza A virus is an enveloped virus that contains two very important surface glycoproteins: H and N. The H and N genes contain the hot spots that code for antigenic sites recognized by virus-neutralizing antibodies. Virus-neutralizing antibodies defend host cells from viral attack by inhibiting the effects of the virus during infection. Epidemics and pandemics occur when there are changes in the antigenic structure of the H and N glycoproteins of the influenza A virus. Two processes—antigenic drift and antigenic shift—mediate the genetic variation, giving rise to new strains of influenza A virus (FIGURE 9-13).

FIGURE 9-13 (a) Antigenic drift illustration at the cellular level. During replication of the influenza A or B virus genomic RNA, mutation(s) occur. Antigenic drift is the gradual accumulation of new antigenic changes of the H protein (and, to a lesser degree, the N protein). (b) Antigenic drift is occurring all of the time among influenza viruses that infect human populations.
Antigenic Drift
Antigenic drift is the result of mutations in the H and/or N gene that cause small changes in the hemagglutinin and neuraminidase glycoproteins that happen continually over time as influenza A virus replicates. New virus strains are produced that may not be recognized by the body’s immune system, necessitating changes in influenza vaccines. When a person is infected with a particular influenza virus strain, the individual develops antibodies against that virus. As newer virus strains appear, the antibodies against the older strains no longer recognize the newer virus, and reinfection can occur. This is one of the main reasons why people can get influenza more than once. Antigenic drift is responsible for localized and/or seasonal influenza epidemics caused by influenza A or B virus. In most years, one or two of the three virus strains used to produce influenza vaccine are updated with the changes in the circulating influenza viruses (Figure 9-13).
Antigenic shift
Antigenic shift occurs when two different influenza A viruses—one of nonhuman origin (usually from wild water-fowl) and the other of human origin—both infect the same epithelial cell present within the lining of the respiratory tract of a pig or bird, resulting in a novel influenza A virus (FIGURE 9-14). This progeny “hybrid” virus may contain major changes in the H and N genes that are so different from subtypes circulating in humans that people do not have immunity to it. When antigenic shifts occur, herd immunity does not exist. Large numbers of people do not have antibodies against the novel influenza A virus and are susceptible to infection, leading to an influenza pandemic.
Antigenic shifts result in an unusually high number of influenza cases and deaths for approximately 2–3 years. Influenza A viruses infect mammals, including humans, and birds. Antigenic shift does not occur with influenza B viruses, because influenza B viruses can only infect humans (Table 9-2).
Normally, avian influenza A viruses replicate poorly in humans. Conversely, human influenza A viruses replicate poorly in birds. Reassortments between human and bird influenza A viruses may produce a new subtype of influenza A virus that infects and replicates well in humans. The virus is able to reassort its genes, swapping RNA segments of the virus genome in coinfected host cells. Reassortment of influenza A viruses results in new H#N# strains plus 256 other possible influenza A virus genomic combinations.
FIGURE 9-15 is an illustration showing the different reservoirs of influenza A viruses. Wild waterfowl (e.g., ducks and geese) are the largest reservoir of influenza A viruses. Infections in wild waterfowl and shorebirds are usually asymptomatic. Influenza A viruses replicate in the intestinal tract and are transmitted by an oral–fecal route. The evolution (i.e., mutation rate) of influenza A virus genomes in wild aquatic birds is very low. However, upon transmission to domesticated birds (e.g., chickens or turkeys), rapid evolution occurs.
Replication of influenza A viruses of H5 or H7 sub-types can lead to the emergence of highly pathogenic influenza A viruses that cause devastating epizootics. An epizootic is an epidemic among animals or birds of a single population within a particular region. This happened during the fall of 2014 when a highly pathogenic influenza A virus (H5N2) was identified in commercial poultry in southern British Columbia, Canada. Surveillance activities determined that the highly pathogenic influenza A virus was found in wild birds in the United States. Surveillance activities targeting wild bird samples from hunter-killed ducks or geese determined that every state in the Pacific flyway had at least one sample that tested positive for highly pathogenic influenza A viruses. Further screening found highly pathogenic influenza A viruses in additional flyways, along with multiple epizootics in backyard and commercial poultry.
Migratory Birds and the Spread of Highly Pathogenic Influenza a Viruses
Many species of birds migrate long distances between their breeding grounds and nonbreeding locations. As natural reservoirs of influenza A viruses, waterfowl and shorebirds play an important role in the maintenance and spread of low-pathogenic influenza A viruses and possibly in the spread of highly pathogenic influenza A viruses such as the H5N1 and H5N2 subtypes. Migration patterns are well documented.
The extent of migratory movements can vary great both among and within species. Migratory routes of birds are grouped together as flyways. A flyway is the entire range of a migratory bird species through which it travels on an annual basis from the breeding to nonbreeding grounds, including intermediate resting and feeding places. The Pacific flyway is a major north–south flyway for migratory birds in North America, extending from Alaska to Patagonia. Every year migratory birds travel in the spring and fall, following food sources to breeding grounds or overwintering sites (FIGURE 9-16). The birds traverse diverse habitats, from Arctic tundra and northwestern rainforest to tropical beaches and man-groves. Within North America, there are four flyways: Atlantic, Mississippi, Central, and Pacific. The Pacific flyway is thought to be the most likely area of introduction for highly pathogenic influenza A viruses in the United States and Canada (FIGURE 9-17).
The Swine-Origin Influenza a Virus (H1N1)
The epicenter of the 2009 influenza pandemic was the Mexican town of La Gloria, Veracruz. The first cases occurred in mid-February of 2009. By early April, Mexican public health officials were investigating high numbers of pneumonia/influenza-like illnesses. They informed the Pan American Health Organization (PAHO) and World Health Organization (WHO) of a possible pandemic. In April, CDC experts collected two clinical specimens from patients in southern California. The respiratory viruses were isolated from patient specimens, and further investigation revealed that the Mexican and Californian cases were caused by similar influenza A viruses. Data on the genetic composition of the influenza A virus isolates became available shortly after virus isolation. It was determined to be an H1N1 subtype. The CDC experts developed an RT-PCR (reverse-transcriptase polymerase chain reaction) diagnostic test kit to detect the 2009 influenza A virus (H1N1) pandemic strain.

FIGURE 9-14 (a) (a) Antigenic shift illustration at the cellular level. Antigenic shift occurs when influenza A viruses acquire a new H or N gene, resulting in a new strain or subtype. (b) Antigenic shift illustration at the host level. Antigenic shifts allow influenza A viruses to move from animals or birds to infect humans. Large numbers of individuals do not possess antibodies to protect them from infection by the novel influenza A virus.

FIGURE 9-15 The reservoir and interspecies transmission of influenza A viruses. Wild waterfowl and shorebirds, domestic birds, pigs, horses, humans, and bats maintain their own influenza A viruses, indicated by circular arrows and subtypes in red. Spillover events occur occasionally, usually from wild birds, indicated by the straight arrow and subtypes in green. Influenza A virus (H7N7) emerged among horses in the 1950s but is currently thought to be extinct.

FIGURE 9-16 Flyways of migratory birds. Some species of birds have very long transequatorial annual migrations, breeding in the high latitudes of the Arctic during the northern summer and then traveling to more hospitable middle or southern latitudes as far south as South America. Some migratory species spread low pathogenic strains of avian influenza A viruses and potentially spread highly pathogenic influenza A viruses.

FIGURE 9-17 North America has four main flyways that represent the migratory paths of many species: Pacific, Central, Mississippi, and Atlantic. The four flyways have areas that tend to overlap and converge. The flyway boundaries are defined based on known bird migration routes but are not geographically fixed or sharply defined.
The influenza A virus (H1N1) was alarming to experts because it was genetically traced to an antigenic shift that resulted in a triple reassortment virus of avian, human, and swine origin: North American avian influenza A virus + classical swine influenza A virus + a human influenza A virus (H3N2) that was circulating in the 1990s. Subsequently, a Eurasian avian-like swine influenza A virus circulated within the classical pig population, resulting in a novel reassortment influenza A virus that infected humans. The final pandemic human influenza A virus (H1N1) contained genes of the following origins: PB2-North American avian, PB1-human H2N2, PA-North American avian, H1-classical swine, NP-classical swine, N1-Eurasian avian-like swine, and NS-classical swine (FIGURE 9-18). A timeline showing the relationship between pandemic influenza A virus strains between 1900 and 2010 is shown in FIGURE 9-19.
Is China the Incubator for Influenza A Viruses?
Why does it seem that pandemic influenza A virus strains originate in southern China? Some suggest that this is because there is a close association of humans with domestic animals and birds raised for food (FIGURE 9-20). Influenza A viruses are constantly circulating in humans, wild birds, and animals (Figure 9-15). There are many small farms in China and other countries in Southeast Asia. The likelihood of genetic reassortment of influenza A viruses between humans and other species is considerable. In addition to extensive small-scale farming, China and Southeast Asian countries are densely populated. This may be a factor in the evolution of influenza A viruses. Of course, these are hypotheses; there is no proof that epidemics originate this way.
Why Is Influenza More Prevalent During the Winter?
In temperate climates such as North America, influenza generally affects people from November to March, whereas influenza A virus infections occur throughout the year in tropical climates. A possible explanation for the high level of influenza activity during the winter is that the gathering of people indoors facilitates the transmission of influenza A viruses. A 2007 study by Anice Lowen and Peter Palese’s research teams demonstrated that the aerosol spread of influenza A virus was dependent upon humidity and temperature. Influenza A viruses were best transmitted at low humidity (20%) and colder temperatures (41°F [5°C]). They provided supporting evidence that environmental conditions play a role in influenza A virus transmission. Findings from a 2010 study confirmed that humidity drives seasonal variation of influenza A virus transmission in temperate regions.
9.10 Influenza Pandemics in History
During the 20th century, three influenza A pandemics occurred: the Spanish influenza of 1918–1919, the Asian influenza of 1957–1958, and the Hong Kong influenza of 1968–1969. TABLE 9-7 lists the U.S. mortalities and influenza A virus subtypes associated with each pandemic.

FIGURE 9-18 Genesis of the 2009 swine influenza A virus (H1N1). A triple reassortment between classical swine influenza A viruses, human influenza A virus (H3N2), and North American avian influenza A viruses occurred sometime during the late 1990s. Reassortments of swine influenza A virus circulated in North American pig populations. A classical swine influenza A virus triple reassortment underwent a subsequent reassortment with a Eurasian avian-like swine influenza A virus, resulting in a swine influenza A virus (H1N1) that circulated in humans in 2009.

FIGURE 9-19 Timeline showing the relationship of H1N1 human and swine influenza A viruses. The 1918 influenza A pandemic virus gave rise to influenza A viruses (H1N1), causing seasonal epidemics that disappeared in 1957 and reemerged in 1977. A seasonal 1946 influenza epidemic affecting soldiers at Fort Dix (United States) caused by a classical swine influenza A virus in 1976 is also indicated on the timeline. During the late 1990s, the triple reassortment influenza A virus emerged in North American pigs, which contained PB2 and PA of a North American avian influenza A virus, PB1 and N of a human influenza A virus (H3N2), and the other four genes of classical North American swine influenza A virus. The 2009 novel/pandemic influenza A virus (H1N1) is a triple reassortment with the N and M genes from the Eurasian avian-like H1N1 lineage that emerged around 1970 in North American pigs.
The 1918 Spanish Influenza
The 1918 Spanish influenza killed more people in 25 weeks than AIDS killed in 25 years. It killed more people in a year than the plagues of the Middle Ages killed in a century. Estimated deaths range between 20 and 50 million, including 675,000 Americans. In New York City alone its impact resulted in 21,000 children orphaned after their parents died from influenza. Seven times as many people died of the 1918 influenza than died in World War I (WWI). True mortality numbers will never be known because many places were overwhelmed by influenza cases and did not keep mortality statistics. The 1918 pandemic spread faster than any plague in history, charging across America in a mere 7 days. It took only 3 months to sweep around the world. In Alaska, 60% of the Inuit population was wiped out by influenza. Islands in the South Pacific, a region where respiratory illness is uncommon and not life-threatening, lost 20% of their population. The 1918 Spanish influenza was 25 times more deadly than previous influenza A viruses.

FIGURE 9-20 (a) Farmers migrating in China bring birds and animals with them. Humans working in close association with birds and mammals facilitate the evolution and genetic variation of influenza A viruses. (b) It is not uncommon for farmers in Southeast Asia to live with their birds and animals; for example, these chickens live with a family that works on a small farm.
Table 9-7 Influenza Pandemics in History
| Dates | U.S. Deaths | Influenza Strain |
|---|---|---|
| 1918–1919 “Spanish flu” | 675,000 (> 20 million worldwide) | H1N1 |
| 1957–1958 “Asian flu” | 70,000 | H2N2 A/Singapore/57 |
| 1968–1969 “Hong Kong flu” | 33,800 | H3N2 A/Aichi/68 |
| 2009 | 7,500–12,000* | H1N1 A/Mexico/09 |
*Conservative range estimate based on CDC survey. Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
 Get Clinical Tree app for offline access
Get Clinical Tree app for offline access

| ||