Chapter 2 Inflammation and Repair



ACUTE INFLAMMATION
5 What are the three main sets of events taking place in tissues during acute inflammation?
Three main events in acute inflammation are:



6 What is the difference between an exudate and a transudate?
TABLE 2-1 Differences Between Transudate and Exudate
| Transudate | Exudate | |
|---|---|---|
| Appearance | Clear | Turbid |
| Specific gravity | <1.015 | >1.020 |
| Protein content | <3 g/dL | >3 g/dL |
| Cells | Scant | Numerous neutrophils |
8 Why does edema develop in acute inflammation?
Inflammatory edema has three main causes:
 Increased hydrostatic pressure in microcirculation: Arteriolar dilatation associated with an increased influx of blood promotes transudation of fluid in capillaries and venules.
Increased hydrostatic pressure in microcirculation: Arteriolar dilatation associated with an increased influx of blood promotes transudation of fluid in capillaries and venules.

9 How does the permeability of small vessels increase during inflammation?
There are several mechanisms, the most important of which are the following:
 Formation of gaps between endothelial cells: This is the most common form of increased permeability. In early stages of inflammation, it occurs predominantly in venules under the influence of histamine or bradykinin. In later stages of inflammation, it is mediated by cytokines. These mediators of inflammation cause widening of intercellular gaps as a result of retraction of endothelial cells.
Formation of gaps between endothelial cells: This is the most common form of increased permeability. In early stages of inflammation, it occurs predominantly in venules under the influence of histamine or bradykinin. In later stages of inflammation, it is mediated by cytokines. These mediators of inflammation cause widening of intercellular gaps as a result of retraction of endothelial cells.


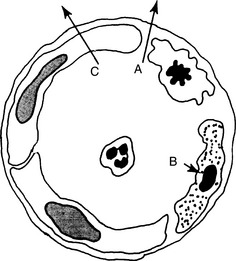
10 What are the main events leading to transmigration of leukocytes across the vessel wall?
 Margination: The slowing of the blood flow allows the neutrophils to exit from the center of the bloodstream into the peripheral part and thus establish contact with endothelial cells.
Margination: The slowing of the blood flow allows the neutrophils to exit from the center of the bloodstream into the peripheral part and thus establish contact with endothelial cells.


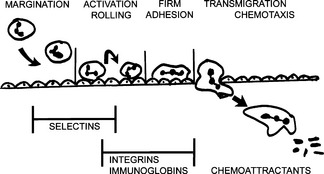
13 What are the signs of leukocyte activation?
Activated leukocytes differ from inactive leukocytes in several respects:
 Expression of adhesion molecules: Selectins and integrins appear on the cell surface or are expressed in higher numbers and show higher affinity for liquids.
Expression of adhesion molecules: Selectins and integrins appear on the cell surface or are expressed in higher numbers and show higher affinity for liquids.



14 What is chemotaxis?
Chemotaxis is active movement of cells along a chemical gradient generated by a chemoattractant.







18 How do leukocytes kill bacteria?
The killing of bacteria occurs in three stages and involves:



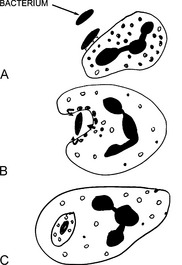



22 What are the main bactericidal substances used by neutrophils?
Bacteria can be killed through two mechanisms:
 Oxygen-dependent killing: This mechanism depends on the oxygen burst resulting from the activation of nicotinamide adenine dinucleotide phosphate (NADPH) oxidase. Oxidation of NADPH generates superoxide, which spontaneously transforms into hydrogen peroxide (H2O2). H2O2 is the substrate for myeloperoxidase, which links it to a chloride ion, generating hypochloric acid. Hypochloric acid, similar to household Clorox, is the most potent bactericidal chemical generated in the phagosomes.
Oxygen-dependent killing: This mechanism depends on the oxygen burst resulting from the activation of nicotinamide adenine dinucleotide phosphate (NADPH) oxidase. Oxidation of NADPH generates superoxide, which spontaneously transforms into hydrogen peroxide (H2O2). H2O2 is the substrate for myeloperoxidase, which links it to a chloride ion, generating hypochloric acid. Hypochloric acid, similar to household Clorox, is the most potent bactericidal chemical generated in the phagosomes.
24 What are the most important congenital defects of leukocyte function?
 Defects in leukocyte adhesion: This category encompasses deficiencies of various leukocyte adhesion molecules, integrins, and enzymes that synthesize the carbohydrate ligands for the selectins.
Defects in leukocyte adhesion: This category encompasses deficiencies of various leukocyte adhesion molecules, integrins, and enzymes that synthesize the carbohydrate ligands for the selectins.

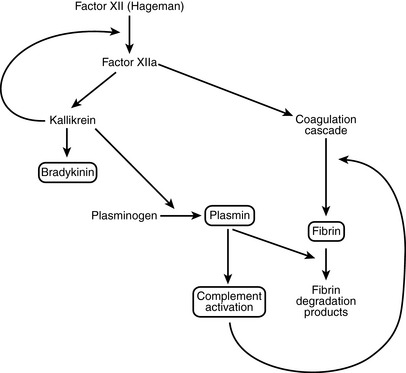
25 What are the most important plasma-derived mediators of inflammation?
 Kinin system: This results in the formation of kallikrein and bradykinin. Kallikrein itself is capable of activating Hageman factor and could play a role in autocatalytic propagation of the entire enzymatic cascade.
Kinin system: This results in the formation of kallikrein and bradykinin. Kallikrein itself is capable of activating Hageman factor and could play a role in autocatalytic propagation of the entire enzymatic cascade.


< div class='tao-gold-member'>
Only gold members can continue reading. Log In or Register to continue
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


