found in virtually all forms of life. Tetrahydrofolate (THF), which is the fully reduced form of the vitamin, carries one-carbons at one of three different oxidation levels ranging from methanol to formate (5, 6). The one-carbons are covalently bound to the N5 or N10 position of THF. In the cell, five different one-carbon substituted forms of THF are present: 10-formyl-THF; 5-formyl-THF; 5,10-methenyl-THF; 5,10-methylene-THF; and 5-methyl-THF, and each of these forms is interconverted in the cell through enzyme-mediated catalysis. Folates are also modified through the addition of a glutamate polypeptide that is polymerized through unusual γ-linked peptide bonds (7). The polyglutamate polypeptide increases the affinity of folate cofactors for folate-dependent enzymes and is required to retain folates within the cell and subcellular organelles. Folic acid (see Fig. 26.1) is not a biologically active form of folate, but it can serve as a provitamin because it is converted to the reduced, natural form of folate once transported into cells. It is an oxidized form of folate generated during the oxidative degradation of folate and normally does not accumulate in cells, although most degradation of THF is irreversible with degradation products that include oxidized pterin and para-aminobenzoyl-glutamate (8). Folic acid is also a synthetic form of folate present in fortified foods and in dietary supplements.
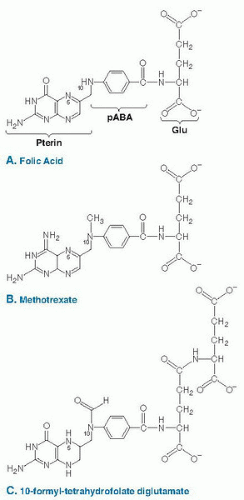 Fig. 26.1. The chemical structure of folic acid (A), methotrexate (B), and 10-formyl-tetrahydrofolate diglutamate (C). Folic acid contains a pterin ring that is bridged to para-aminobenzoic acid (PABA) through a methylene group to form pteroic acid. The addition of the glutamate residue (Glu) through a peptide linkage results in the formation of folic acid. Methotrexate (4-amino-10-methylpteroylglutamic acid) (B) is a folate analog, antagonist, and pharmaceutical agent that inhibits the activity of DHFR. Once transported into the cell, folic acid is reduced to tetrahydrofolate and is modified by the addition of a glutamate polypeptide containing up to nine glutamate residues linked by unusual γ-peptide linkages. THF is also modified by the addition of single carbons at the N5 or N10 position or that bridge the N5 and N10 positions. The carbon moieties are carried at the oxidation states of formate, formaldehyde, or methanol. The structure of 10-formyl-tetrahydrofolate diglutamate is shown in C. |
TABLE 26.1 DIETARY REFERENCE INTAKES FOR FOLATEa | ||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||
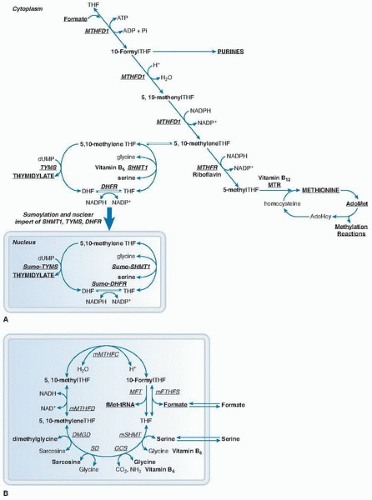 Fig. 26.2. Compartmentation of folate-mediated one-carbon metabolism in the cytoplasm, mitochondria, and nucleus. A. One-carbon metabolism in the cytoplasm is required for the de novo synthesis of purines and thymidylate and for the remethylation of homocysteine to methionine. One-carbon metabolism in the nucleus synthesizes thymidylate from uridylate and serine, and it occurs during the S phase of the cell cycle. B. One-carbon metabolism in the mitochondria is required to generate formate for one-carbon metabolism in the cytoplasm. The folate and amino acid carriers of the one-carbon unit are indicated by bold type. AdoHcy, S-adenosylhomocysteine; AdoMet, S-adenosylmethionine; ADP, adenosine diphosphate; ATP, adenosine triphosphate; DHF, dihydrofolate; DHFR, dihydrofolate reductase; DMGD, dimethylglycine dehydrogenase; dUMP, deoxyuridine monophosphate; GCS, glycine cleavage system; mFTHFS, mitochondrial formyltetrahydrofolate synthetase; mMTHFC, mitochondrial methenyltetrahydrofolate cyclohydrolase; mMTHFD, mitochondrial methylenetetrahydrofolate dehydrogenase; MTHFR, methylenetetrahydrofolate reductase; MTR, methionine synthase; M-tRNA-FT, methionyl-tRNA formyltransferase; NAD, nicotinamide adenine dinucleotide; NADP, nicotinamide adenine dinucleotide phosphate; NADPH, reduced nicotinamide adenine dinucleotide phosphate; Pi, inorganic phosphate; SD, sarcosine dehydrogenase; SHMT1, cytoplasmic serine hydroxymethyltransferase; THF, tetrahydrofolate; TYMS, thymidylate synthase. |
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


