CHAPTER 2 Evolution of Life on Earth
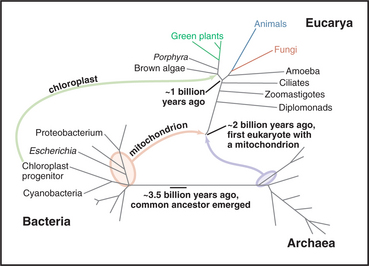
No one is certain how life began, but the common ancestor of all living things populated the earth over 3 billion years ago, not long (geologically speaking) after the planet formed 4.5 billion years ago (Fig. 2-1). Biochemical features shared by all existing cells suggest that this primitive microscopic cell had about 600 genes encoded in DNA, ribosomes to synthesize proteins, and a plasma membrane with pumps, carriers, and channels. Over time, mutations in the DNA created progeny that diverged genetically into numerous distinctive species, numbering about 1.7 million known to science. The total number of species living on the earth today is unknown but is estimated to be between 4 million and 100 million. On the basis of evolutionary histories preserved in their genomes, living organisms are divided into three primary domains: Bacteria, Archaea, and Eucarya.
Evolution is the great unifying principle in biology. Research on evolution is both exciting and challenging because this ultimate detective story involves piecing together fragmentary evidence spread over 3.5 billion years. Data include fossils of ancient organisms preserved in stone, ancient DNA (going back about 45,000 years), and especially DNA of living organisms.
Prebiotic Chemistry Leading to an RNA World
But where did the common ancestor come from? A wide range of evidence supports the idea that life began with self-replicating RNA polymers sheltered inside lipid vesicles even before the invention of protein synthesis (Fig. 2-2). This hypothetical early stage of evolution is called the RNA World. This postulate is attractive because it solves the chicken-and-egg problem of how to build a system of self-replicating molecules without having to invent either DNA or proteins on their own. Clearly, RNA has an advantage, because it provides a way to store information in a type of molecule that can also have catalytic activity. Proteins excel in catalysis but do not store self-replicating genetic information. Today, proteins have largely superseded RNAs as cellular catalysts. DNA excels for storing genetic information, since the absence of the 2′ hydroxyl makes it less reactive and therefore more stable than RNA. Readers who are not familiar with the structure of nucleic acids should consult Chapter 3 at this point.
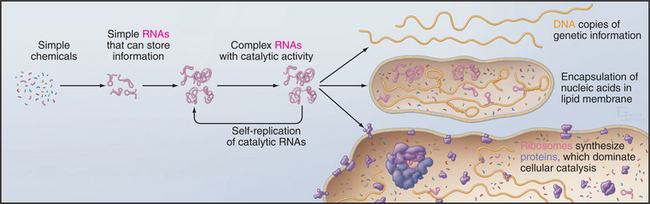
Given a supply of nucleotides, these reactions could have created a heterogeneous pool of small RNAs, the biochemical materials required to set in motion the process of natural selection at the molecular level. The idea is that random sequences of RNA are selected for replication on the basis of useful attributes. This process of molecular evolution can now be reproduced in the laboratory by using multiple rounds of error-prone replication of RNA to produce variants from a pool of random initial sequences. Given a laboratory assay for a particular function, it is possible to use this process of directed evolution to select RNAs that are capable of catalyzing biochemical reactions (called ribozymes), including RNA-dependent synthesis of a complementary RNA strand. Although unlikely, this is presumed to have occurred in nature, creating a reliable mechanism to replicate RNAs. Subsequent errors in replication produced variant RNAs, some having desirable features such as catalytic activities that were required for a self-replicating system. Over millions of years, a ribozyme eventually evolved with the ability to catalyze the formation of peptide bonds and to synthesize proteins. This most complicated of all known ribozymes is, of course, the ribosome (see Fig. 17-6) that catalyzes the synthesis of proteins. Proteins eventually supplanted ribozymes as catalysts for most biochemical reactions. Owing to greater chemical stability, DNA proved to be superior to RNA for storing the genetic blueprint over time.
Each of these events is improbable, and their combined probability is exceedingly remote, but given a vast number of chemical “experiments” over hundreds of millions of years, this all happened. Encapsulation of these prebiotic reactions may have enhanced their probability. In addition to catalyzing RNA synthesis, clay minerals can also promote formation of lipid vesicles, which can corral reactants to avoid dilution and loss of valuable constituents. This process might have started with fragile bilayers of fatty acids that were later supplanted by more robust phosphoglyceride bilayers (see Fig. 7-5). In laboratory experiments, RNAs inside lipid vesicles can create osmotic pressure that favors expansion of the bilayer at the expense of vesicles lacking RNAs.
Another mystery is how l-amino acids and d-sugars (see Chapter 3) were selected over their stereoisomers for biomacromolecules. This was a pivotal event, since racemic mixtures are not favorable for biosynthesis. For example, mixtures of nucleotides composed of l- and d-ribose cannot base-pair well enough for template-guided replication of nucleic acids. In the laboratory, particular amino acid stereoisomers (that could have come from meteorites) can bias the synthesis of D-sugars.
Divergent Evolution from the Last Universal Common Ancestor of Life
Shared biochemical features suggest that all current cells are derived from a last universal common ances-tor about 3.5 billion years ago (Fig. 2-1). This primitive ancestor could, literally, have been a single cell or colony of cells, but it might have been a larger community of cells sharing a common pool of genes through interchange of their nucleic acids. The situation is obscure because no primitive organisms remain. All contemporary organisms have diverged equally far in time from their common ancestor.
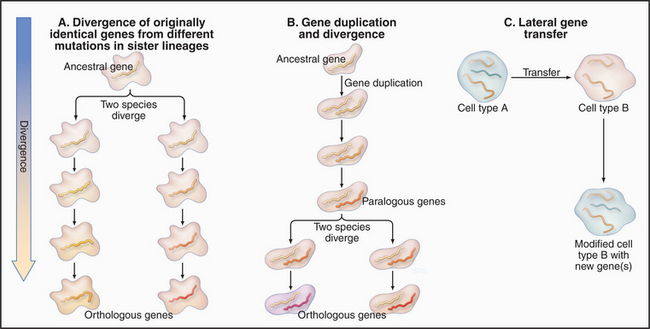
During evolution genomes have diversified by three processes (Fig. 2-3):
Evolution of Prokaryotes
Since the beginning of life, microorganisms dominated the earth in terms of numbers, variety of species, and range of habitats (Fig. 2-4). Bacteria and Archaea remain the most abundant organisms in the seas and on land. They share many features, including basic metabolic enzymes and flagella powered by rotary motors embedded in the plasma membrane. Both divisions of prokaryotes are diverse with respect to size, shape, nutrient sources, and environmental tolerances, so these features cannot be used for classification, which relies instead on analysis of their genomes. For example, sequences of the genes for ribosomal RNAs cleanly separate Bacteria and Archaea (Fig. 2-4). Bacteria are also distinguished by plasma membranes of phosphoglycerides (see Fig. 7-5
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


