2 EPITHELIAL GLANDS
Development of epithelial glands
Most glands develop as epithelial outgrowths into the underlying connective tissue (Figure 2-1). Exocrine glands remain connected to the surface of the epithelium by an excretory duct that transports the secretory product to the outside. Endocrine glands lack an excretory duct, and their product is released into the blood circulation.
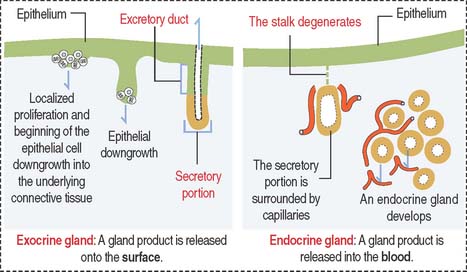
Endocrine glands are surrounded by fenestrated capillaries and commonly store the secretions they synthesize and release after stimulation by chemical or electrical signals. Exocrine and endocrine glands can be found together (for example, in the pancreas), as separate structures in endocrine organs (thyroid, parathyroid), or as single cells (enteroendocrine cells). Endocrine glands will be studied later in Chapter 18, Neuroendocrine System, and Chapter 19, Endocrine System.
Classification of epithelial glands
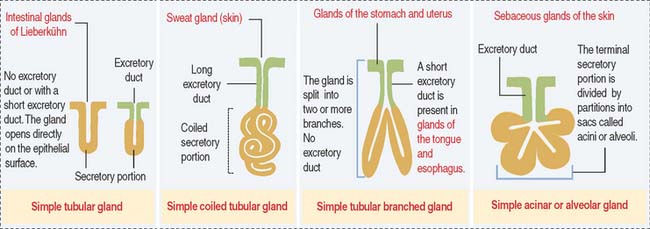
Glands are classified according to the type of excretory duct into simple and branched (also called compound) glands. The gland can be simple (Figure 2-2) when the excretory duct is unbranched. The gland can be branched when the excretory duct subdivides (Figure 2-3).
The secretory portion can be unicellular or multicellular
An exocrine gland has two components: a secretory portion and an excretory duct. The secretory portion of a gland may be composed of one cell type (unicellular, for example, goblet cells in the respiratory epithelium and intestine) or many cells (multicellular).
According to the shape of the secretory portion (see Figures 2-2 and 2-3), glands can be tubular, coiled, or alveolar (Latin alveolus, small hollow sac; plural alveoli), also called acinar (Latin acinus, grape; plural acini).
Shape of the secretory portion
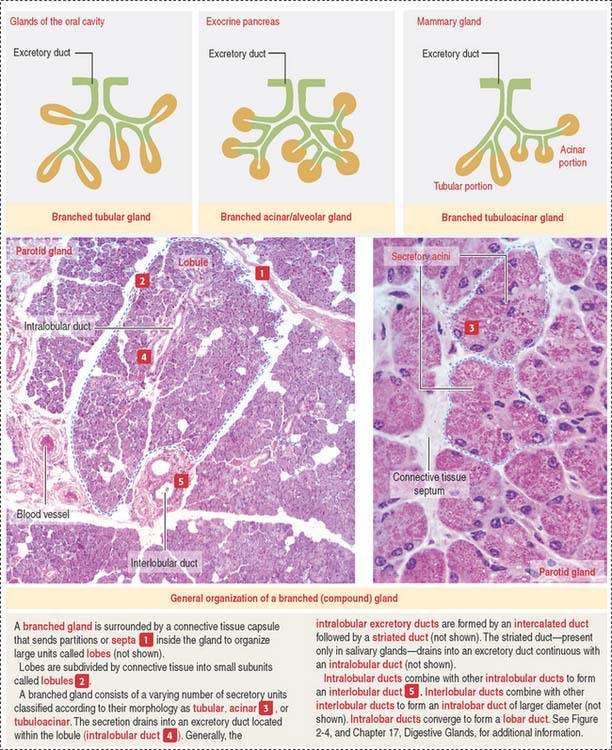
A branched gland (Figure 2-4) is surrounded by a capsule. Septa or trabeculae extend from the capsule into the glandular tissue. Large septa divide the gland into a number of lobes. Branches from the septa separating adjacent lobes divide the lobes into smaller compartments called lobules.
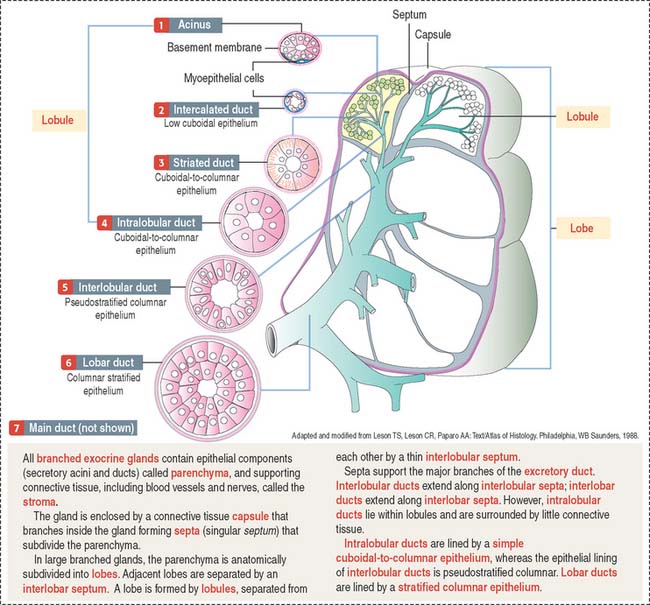
During development, a main excretory duct gives rise to branches that lie either between (interlobar) or within lobes (intralobar). Small branches derived from each of these ducts generate small subdivisions that constitute the lobule of a gland. These branches can be found first between lobules (interlobular) and within lobules (intralobular). Additional details are presented in Chapter 17, Digestive Glands.
Types of secretion
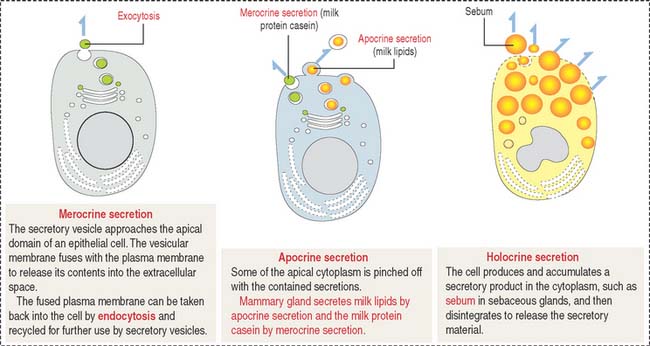
Based on the type of secretion, exocrine glands can be classified as mucous glands, when their products are rich in glycoproteins and water; serous glands, with secretions enriched with proteins and water; and mixed glands, which contain both mucous and serous cells (Figure 2-5).

CYTOMEMBRANES
Plasma membrane
A review of major concepts of cytomembranes and organelles and their clinical relevance is presented in this chapter. Epithelial glands are a convenient topic for this integration. We initiate the review by addressing the structural and biochemical characteristics of the plasma membrane. Additional information related to plasma membrane–mediated cell signaling is presented in Chapter 3, Cell Signaling.
The plasma membrane determines the structural and functional boundaries of a cell. Intracellular membranes, called cytomembranes, separate diverse cellular processes into compartments known as organelles. The nucleus, mitochondria, peroxisomes, and lysosomes are membrane-bound organelles; lipids and glycogen are not membrane-bound and are known as inclusions.
The plasma membrane consists of both lipids and proteins. The phospholipid bilayer is the fundamental structure of the membrane and forms a bilayer barrier between two aqueous compartments: the extracellular and intracellular compartments. Proteins are embedded within the phospholipid bilayer and carry out specific functions of the plasma membrane such as cell-cell recognition and selective transport of molecules (see Box 2-A).
Box 2-A Lipid rafts
Phospholipid bilayer
The four major phospholipids of plasma membranes are phosphatidylcholine, phosphatidylethanolamine, phosphatidylserine, and sphingomyelin (Figure 2-7). They represent more than one half the lipid of most membranes. A fifth phospholipid, phosphatidylinositol, is localized to the inner leaflet of the plasma membrane.
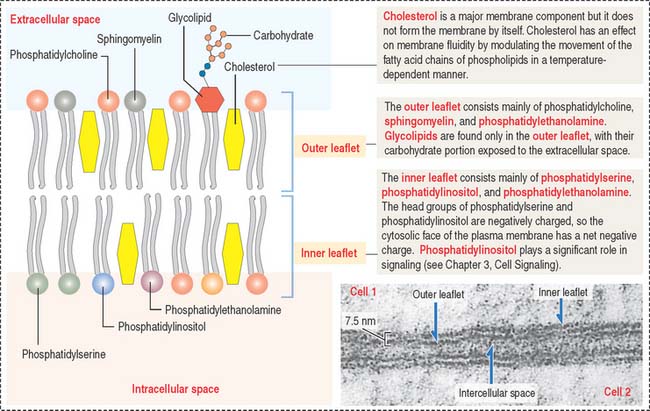
Two general aspects of the phospholipid bilayer are important to remember:
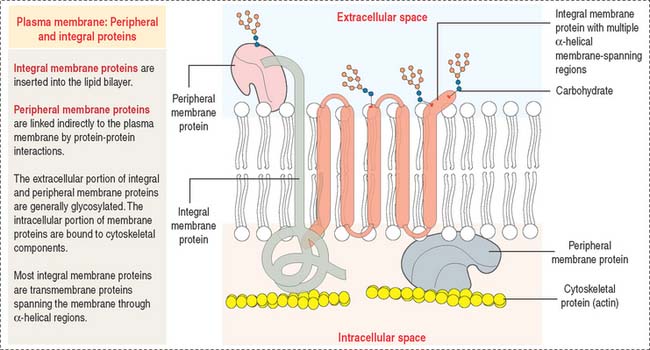
Membrane proteins
Most plasma membranes consist of about 50% lipid and 50% protein (Figure 2-8). The carbohydrate component of glycolipids and glycoproteins represents 5% to 10% of the membrane mass. The surface of a plasma membrane is coated by a glycocalyx (see Box 2-B).
Box 2-B Glycocalyx
Restrictions in the mobility of membrane proteins are responsible for the polarized nature of epithelial cells, divided into distinct apical and basolateral domains that differ in protein composition and function. Tight junctions between adjacent epithelial cells (discussed in Chapter 1, Epithelium) not only seal the space between cells but also serve as barriers to the diffusion of proteins and lipids between the apical and basolateral domains.
Portions of integral membrane proteins are inserted into the lipid bilayer. They can only be released by solubilization using detergents. Detergents are chemical agents that contain both hydrophobic and hydrophilic groups. The hydrophobic domains of the detergent penetrate the membrane lipids and bind to the membrane-inserted hydrophobic portion of the protein. The hydrophilic domains combine with the protein, forming aqueous-soluble detergent-protein complexes.
Freeze-fracture: Differences between a surface and a face
Figure 2-9 indicates the nomenclature for the identification of surfaces and faces in electron micrographs of freeze-fracture preparations.
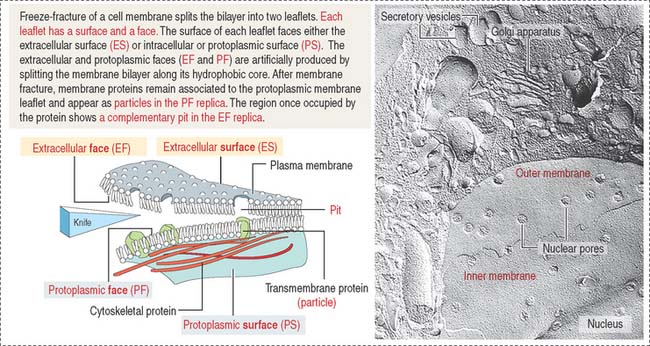
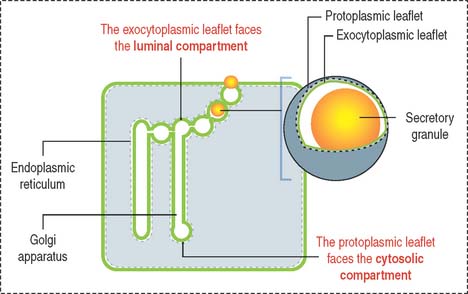
The face of the membrane leaflet looking to the extracellular space (the exocytoplasmic leaflet in the illustration) is labeled EF, for extracellular face. Similarly, the face of the leaflet facing the protoplasmic space (identified as a protoplasmic leaflet) is PF, for protoplasmic face.
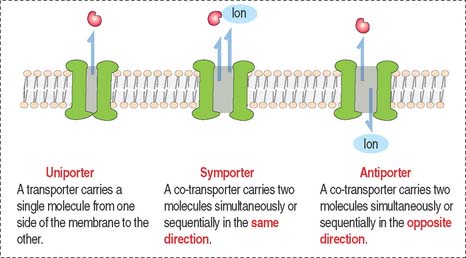
Transporter and channel proteins
Other biological molecules (such as glucose, charged molecules, and small ions—H+, Na+, K+, and Cl−) are unable to dissolve in the hydrophobic interior of the phospholipid bilayer. They require the help of specific transport proteins (Figure 2-10) and channel proteins, which facilitate the diffusion of most biological molecules.
Endoplasmic reticulum
The endoplasmic reticulum is an interconnected network of membrane-bound channels within the cytoplasm, part of the cytomembrane system and distinct from the plasma membrane.
The smooth endoplasmic reticulum lacks ribosomes and is generally in proximity to deposits of glycogen and lipids in the cytoplasm. The smooth endoplasmic reticulum has an important role in detoxification reactions required for the conversion of harmful lipid-soluble or water-insoluble substances into water-soluble compounds more convenient for discharge by the kidneys. It also participates in steroidogenesis (see Chapter 19, Endocrine System).
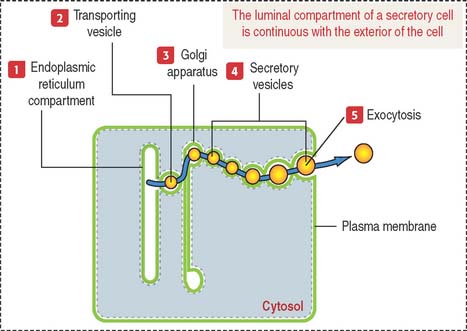
Products released into the luminal compartment of the endoplasmic reticulum are transported to the Golgi apparatus by a transporting vesicle and eventually to the exterior of the cell by exocytosis. One can visualize the sequence in which the lumen of the cytomembrane system is interconnected and remains as such in an imaginary stage; you can visualize that the luminal compartment of a secretory cell is continuous with the exterior of the cell (Figure 2-11). The surrounding space is the cytosolic compartment in which soluble proteins, cytoskeletal components, and organelles are present.
Now, let us visualize the membrane of each component of the cytomembrane system as consisting of two leaflets (Figure 2-12):
Rough endoplasmic reticulum
The rough endoplasmic reticulum is involved in the synthesis of proteins, carried out by their attached ribosomes (Figure 2-13). In contrast, the membranes of the smooth endoplasmic reticulum lack attached ribosomes (see Figure 2-13). Most proteins exit the rough endoplasmic reticulum in vesicles transported to the cis portion of the Golgi apparatus (see Figures 2-16 and 2-17
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


