CHAPTER 20 Endoplasmic Reticulum*
One of the key distinguishing features of eukaryotic cells is the presence of the endoplasmic reticulum (ER), the largest of numerous membrane-delineated intracellular compartments. The ER is thought to have evolved from the prokaryotic plasma membrane by expansion, internalization, and subdivision (Fig. 20-1A-D; see also Fig. 2-6). It provides both an expanded membrane surface (up to 30 times that of the plasma membrane) for carrying out vital cellular functions, including protein and lipid biosynthesis, and an internal compartment (or lumen) that collects proteins synthesized in the cytoplasm for modification and delivery into the secretory pathway. About one third of all cellular proteins are imported into the lumen of the ER or integrated into its membranes. Consisting of an extensive array of tubules or flat saccules called cisternae (cisterna means “reservoir”), the ER forms a continuous three-dimensional network (a reticulum) stretching from the nuclear envelope to the cell surface (Fig. 20-1E). Microtubules and their associated motors generate this extended network in animal cells by pulling ER membranes out toward the periphery of the cell. This allows the ER to coordinate diverse processes over large regions of the cytoplasm. The ER’s size and shape are maintained over time despite a continuous flow of proteins and lipids into and out of this compartment. The flow results from the large rate of synthesis of lipids and proteins (between 2 million and 13 million new proteins per minute) occurring at the ER membrane, as well as from the continuous export of these molecules into the secretory pathway and their selective retrieval back to the ER from the Golgi apparatus.
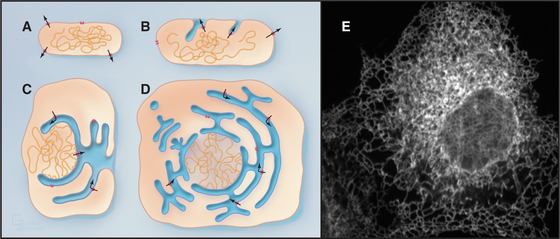
Endoplasmic Reticulum Functions and Organization
The membrane surface of the ER performs several functions for cells. Foremost is the production of the proteins and lipids that will make up the membranes of the other organelles, including the Golgi apparatus, nucleus, endosomes, lysosomes, and plasma membrane, as well as nearly all proteins that will be secreted from the cell. The membranes of mitochondria and peroxisomes also depend on the ER to supply much of their lipid. Another key function of the ER membrane is to form the nuclear envelope, which encloses the nucleus (see Fig. 14-5). The surface of the ER forming the outer nuclear envelope (which faces the cytoplasm) is indistinguishable from the rest of the ER except for the presence of nuclear pores that span both inner and outer nuclear envelope to allow passage of molecules between the nucleus and cytoplasm. By contrast, the ER surface forming the inner nuclear envelope, which faces the nucleus, contains specialized proteins that interact with the nuclear lamina and chromatin. ER membranes also detoxify endogenous steroids, carcinogenic compounds, and lipid-soluble drugs (xenobiotics) from the environment. This occurs by an electron transfer process carried out by ER membrane proteins such as the cytochrome P450 family of enzymes.
The lumen of the ER also performs numerous essential functions. Specialized for receiving proteins transported from the cytoplasm across ER membranes, the ER lumen is enriched in a dense meshwork of chaperones and other modifying enzymes (estimated to be 200 mg/mL in concentration) that catalyze the folding and assembly of newly synthesized proteins. These transported proteins, including both soluble and transmembrane forms, are exported from the ER for secretion or for delivery to the lumen or membrane of the Golgi apparatus, lysosome, or endosome. Proteins that are incorrectly folded or misfolded can be exported back into the cytoplasm, where they are degraded. Misfolded proteins, when accumulated in the ER at high levels, can trigger an unfolded protein response, which activates specific genes in the nucleus whose products help to modify or destroy the misfolded proteins and compensate for the decreased capacity of ER folding. The ER lumen is one of the major Ca2+ storage sites in cells, owing to ER membranes being rich in calcium pumps (see Fig. 8-7) and many Ca2+-binding proteins in the lumen. Such Ca2+ stores can be released by calcium channels in response to cellular signals such as IP3 (see Fig. 26-13). Carefully regulated release and uptake of Ca2+ by the ER control muscle contraction (see Fig. 39-15) and many other cellular processes. The lumen of the ER is also an oxidizing environment that favors disulfide bond formation, which helps to stabilize proteins after they are exported from the ER to the outside of the cell.
The diverse functions carried out by the membrane and in the lumen of the ER (Table 20-1) occur in distinctive ER regions (Fig. 20-2A). The rough ER is studded with ribosomes on its cytoplasmic surface, defining areas that are specialized for protein synthesis, folding, and degradation (Fig. 20-2B). The smooth ER, composed of tubular elements lacking ribosomes, is dedicated to enzyme pathways involved in drug metabolism (hepatocytes), steroid synthesis (endocrine cells), or calcium uptake and release (see Fig. 26-12). The cytochrome P-450 family of heme-containing membrane proteins is found in the smooth ER. Other regions of the ER that lack ribosomes, called ER export domains consist of tubulovesicular membranes that bud during export of secretory cargo to the Golgi apparatus (see Chapter 21). Regions of the ER surrounding the nucleus compose the nuclear envelope.
Table 20-1 SUBDOMAINS OF THE ENDOPLASMIC RETICULUM
| ER Domain | Function | Associated Proteins |
|---|---|---|
| Rough ER | Protein translocation | Sec61 complex, TRAP, TRAM, BiP |
| Protein folding and oligomerization | PDI, Calnexin, Calreticulin, BiP | |
| Carbohydrate addition | Oligosaccharide transferase | |
| ER degradation | EDEM, Derlin1 | |
| Smooth ER | Detoxification | Cytochrome P450 enzymes |
| Lipid metabolism | HMG-CoA reductase | |
| Heme metabolism | Cytochrome b(5) | |
| Calcium release | IP3 receptors | |
| Nuclear envelope | Nuclear pores | POM121, GP210 (see Fig. 14-8) |
| Chromatin anchoring | Lamin B receptor | |
| ER export sites | Export of proteins and lipids into secretory pathway | Sar1p, Sec12p, Sec16p |
| ER contact zones | Transport of lipids | LTPs |
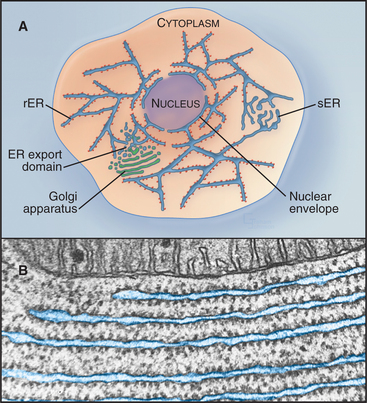
Figure 20-2 endoplasmic reticulum subdomain organization within the cell. A, ER membranes containing ribosomes, called rough ER (rER), extend from the nuclear envelope to the cell periphery. ER membranes lacking ribosomes, called smooth ER (sER), include membranes specialized for drug metabolism and steroid synthesis, as well as tubulovesicular elements composing ER exit sites. The nuclear envelope consists of ER membrane that has wrapped around DNA and other nuclear elements to compartmentalize them. Its double membrane is studded with nuclear pores, which provide a passageway for nuclear components to move between the nucleus and cytosol. (See Fig. 14-17 for more details.) B, Electron micrograph of a thin section of rough ER and neighboring mitochondrion from the pancreas.
(Micrograph by Keith R. Porter; courtesy of Don W. Fawcett, Harvard Medical School, Boston, Massachusetts.)
Overview of Protein Translocation into the Endoplasmic Reticulum
All proteins are synthesized in the cytoplasm and must be specifically targeted to the ER, where they are either fully translocated across the ER membrane and released into the ER lumen (soluble proteins) or only partly translocated across the ER membrane and embedded in the lipid bilayer of the ER membrane (transmembrane proteins). This transport to the ER can happen either as the protein is being made (cotranslational translocation) or after synthesis is complete (posttranslational translocation). The orientation of a protein in the lipid bilayer or its localization to the lumen is established during protein translocation and maintained as the protein is transferred by membrane-bound carriers through the secretory pathway (see Fig. 21-2). Thus, domains of transmembrane proteins to be exposed on the cell surface must be inserted into the ER membrane, facing the lumen. Similarly, secreted soluble proteins must be fully translocated into the lumen of the ER.
Signal Sequence Recognition
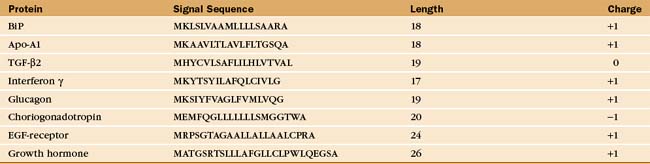
Soluble and membrane proteins destined for ER translocation contain a hydrophobic sequence either at their N-terminus or in transmembrane segments that serve as recognition signals for direction to the ER membrane. N-terminal leader sequences (termed signal sequences) are typically 15 to 35 amino acids long and contain a hydrophobic core of at least 6 residues, while transmembrane signal segments have a hydrophobic stretch of 16 to 25 residues. Aside from hydrophobicity, these signal sequences have no other features in common (Table 20-2). Nevertheless, when attached to proteins that are not normally targeted to the ER, these signal sequences direct the protein to the ER and not to other organelles such as to mitochondria or peroxisomes, which use unique targeting signals (see Fig. 18-1).
Cotranslational Translocation
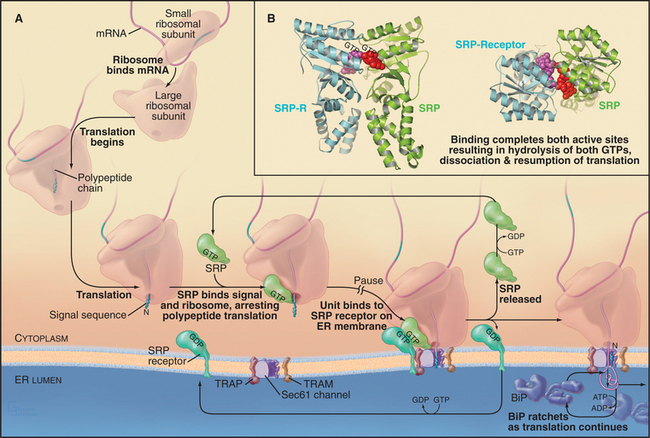
In the cotranslational pathway, the signal recognition particle (SRP) recognizes and binds the first hydrophobic domain of either the signal sequence or the transmembrane signal segment as it emerges from the ribosome slowing translation of the polypeptide (Fig. 20-3A). Then the complex consisting of ribosome, nascent chain, and SRP associates with the ER membrane by binding the SRP receptor (SR), a heterodimer consisting of one subunit that binds SRP and another that spans the ER membrane. Both the SRP and the SR have guanosine triphosphatase (GTPase) domains (Fig. 20-3B) similar to Ras (see Fig. 4-6). GTP binding and hydrolysis by SRP and SR provide directionality and order to the sequence of reactions that bring the nascent chain to the translocation channel. Once the ribosome–nascent chain–SRP complex is bound to SR at the ER membrane, SRP releases the signal sequence, allowing the ribosome–nascent chain complex to be transferred to the protein-conducting channel across the ER membrane. SRP and SR then dissociate after hydrolyzing their bound guanosine triphosphate (GTP), releasing SRP into the cytoplasm and allowing SR to diffuse away in the membrane. The targeting cycle delivers the ribosome–nascent chain complex to the protein-conducting channel (called the translocon) and recycles the targeting machinery (i.e., SRP and SR).
Posttranslational Translocation
Posttranslational translocation (prevalent in fungi such as yeast) differs because neither cotranslational targeting machinery (SRP and SR) nor ribosomes participate. Instead, other components fulfill the functions provided by SRP, SR, and ribosomes (with the Sec61 complex serving as channel, as it does in cotranslational translocation). Signal-containing polypeptides destined for posttranslational insertion into the ER are held in a largely unfolded state by cytoplasmic chaperones until they can be delivered to the tetrameric Sec62/63 protein complex at the ER membrane (Fig. 20-4). A similar strategy is used to import proteins into the mitochondria (see Fig. 18-4). During posttranslational translocation, the signal sequence engages and opens the Sec61 channel in a fashion similar to cotranslational translocation. Since protein synthesis is already complete, another energy source must be exploited to move the substrate through the channel into the ER. BiP, a luminal ER chaperone belonging to the Hsp70 family, binds the substrate in the ER lumen, thereby preventing it from sliding back into the cytoplasm. Repeated rounds of substrate binding and release, catalyzed by ATP hydrolysis, allow BiP to act as a molecular ratchet to drive substrate transport into the lumen. The transmembrane protein Sec63 regulates the ATPase activity of BiP and helps to recruit BiP to the translocation channel. Thus, the peptide is “pulled” across the membrane from the luminal side instead of being “pushed” from the cytoplasmic side, as during cotranslational translocation. Hsp70 family members perform a similar function during import of proteins into mitochondria and chloroplasts (see Fig. 18-4).
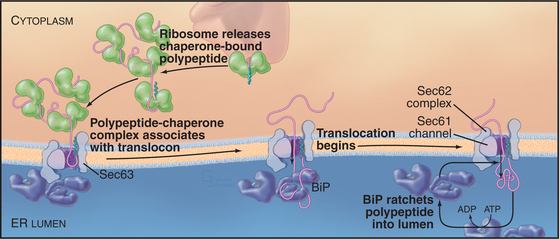
Universality of Protein Translocation
In both cotranslational and posttranslational translocation, the Sec61 complex translocation channel is closed until opened by interacting with binding partners, including the translocated polypeptide. This ensures that only specific types of proteins pass through the ER membrane and that the permeability barrier of these membranes is maintained at all times. In cotranslational translocation, interactions with a ribosome and a signal sequence open the channel, and protein synthesis provides the energy for translocation. In posttranslational translocation, the Sec62/63 complex operates on both sides of the membrane. On the cytoplasmic side, it contributes to the interactions of the signal sequence that open the translocon. On the lumenal side, it recruits BiP to provide the driving force for translocation. This theme applies to protein translocation across the plasma membrane by prokaryotes. There, a homolog of the Sec61 complex (the SecYE complex) interacts with the cytoplasmic SecA adenosine triphosphatase (ATPase), a partner that drives translocation across the plasma membrane (see Fig. 18-9).
Molecular Machinery for Protein Translocation into the Endoplasmic Reticulum
Signal Recognition Particle and Signal Recognition Particle–Receptor
Human SRP is a ribonucleoprotein composed of six proteins (named by their apparent molecular weights and a 300-nucleotide RNA (Fig. 20-5). The SRP54 protein subunit and a portion of the RNA compose the minimal hardware for targeting to the translocon and are found in both prokaryotic and eukaryotic cells. SRP54 binds signal sequences in a deep, hydrophobic groove lined by the flexible side chains of several methionines. Like bristles of a brush, the methionines accommodate the various shapes of the hydrophobic side chains of the residues of different signal sequences. Phosphates of the SRP RNA near one end of this groove may interact with basic residues that are often (but not always) adjacent to the hydrophobic core of signal sequences and transmembrane signal segments.
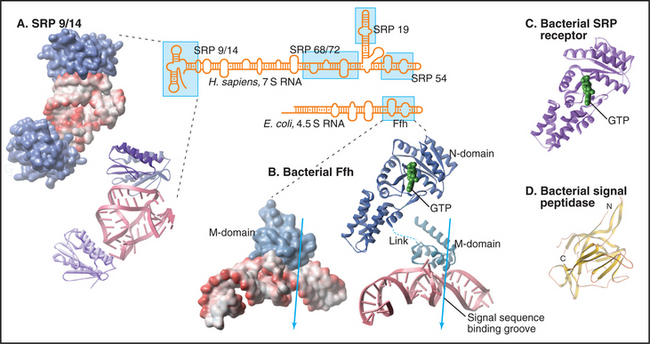
SRP binding to a signal sequence slows translation, a phenomenon that is termed elongation arrest (Fig. 20-3A). The mechanism appears to involve occlusion of the elongation factor binding site on the ribosome by the SRP9 and SRP14 subunits of SRP, which structurally resembles a portion of eEF2. Slowing translation provides time to target the ribosome to the translocation channel before excessive polypeptide synthesis precludes cotranslational transport.
Interaction of SRP with SR directs the SRP–ribosome–nascent chain complex to the translocation channel (Fig. 20-3A). This interaction is regulated by GTP binding and hydrolysis by SRP and SR, whose GTPase cycles are a notable exception to the GTPase switch paradigm of Ras-like GTPases that involve GTP exchange factors (GEFs) and GTPase-activating proteins (GAPs; see Fig. 25-8). No external GEFs or GAPs are known for SRP and SR GTPases. Instead, these proteins readily exchange GDP for GTP and are in the GTP-bound state as they enter the targeting cycle. On formation of a complex, SRP and SR reciprocally activate each other’s GTPase activity, thereby obviating the need for an external GAP to drive the conversion of GTP to GDP on these proteins. Upon GTP hydrolysis, the conformations of SR and SRP change in a way that reduces their affinity. They dissociate from each other as well as from the ribosome–nascent chain and translocation channel. This frees SRP and SR for further rounds of ribosome–nascent chain targeting. Release of SRP and SR occurs only after the ribosome has become properly engaged with the translocation channel in the ER membrane, thereby ensuring that the channel is closed until the ribosome has bound.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


