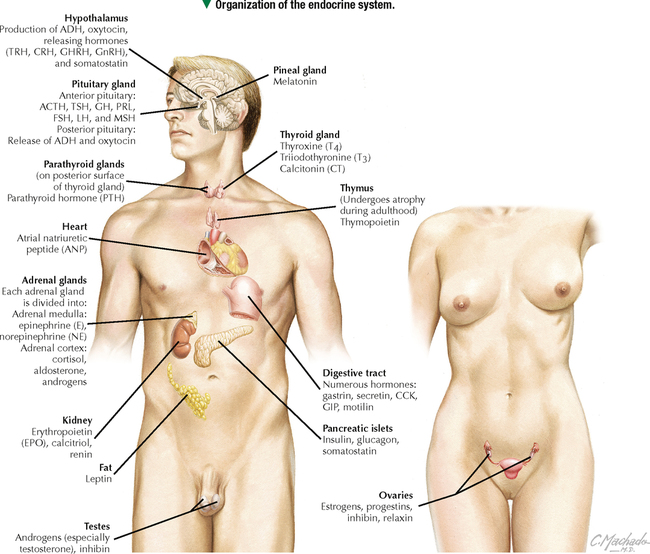
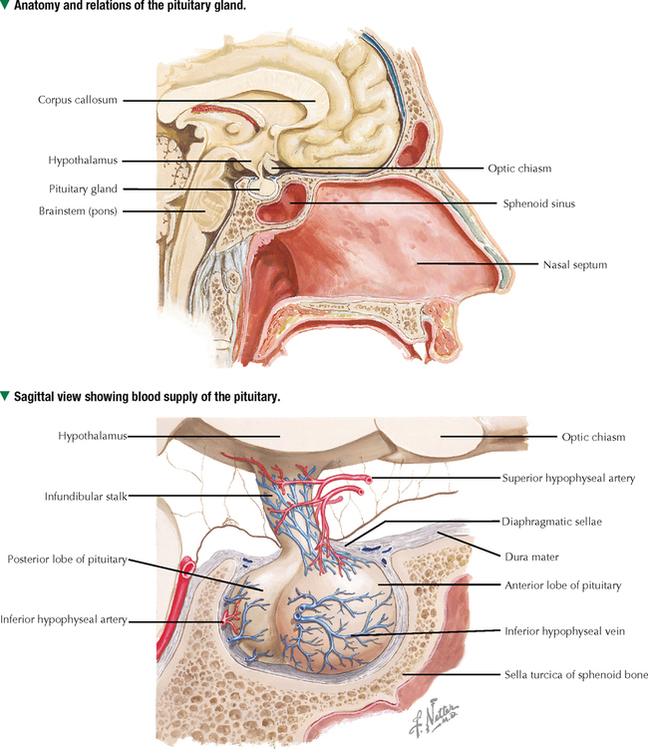
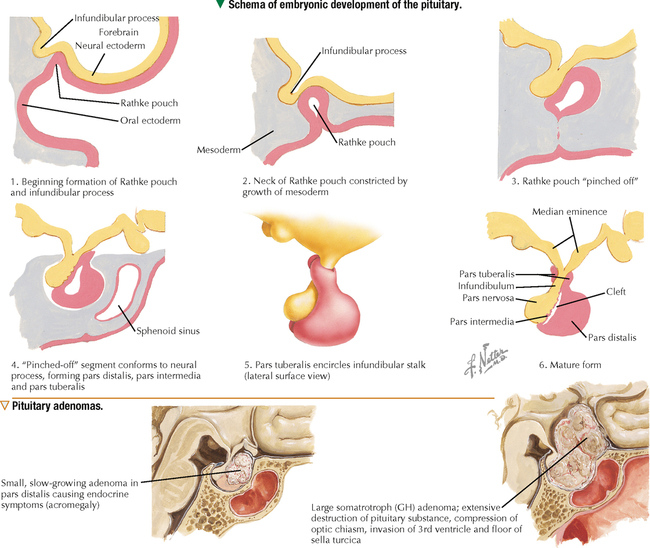
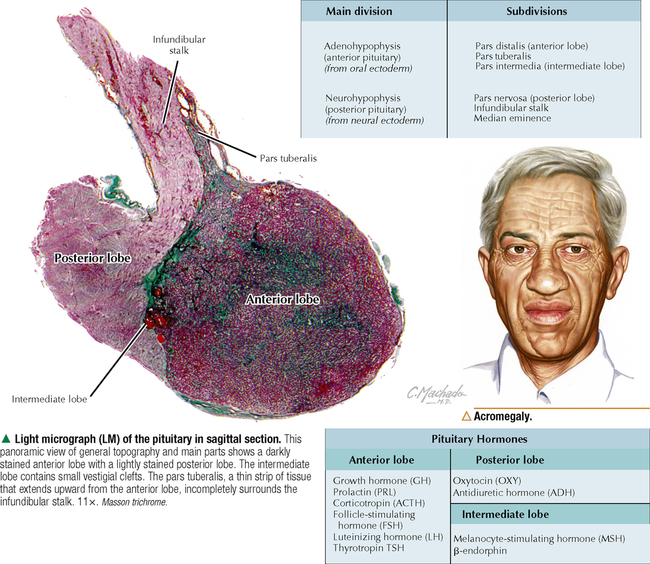
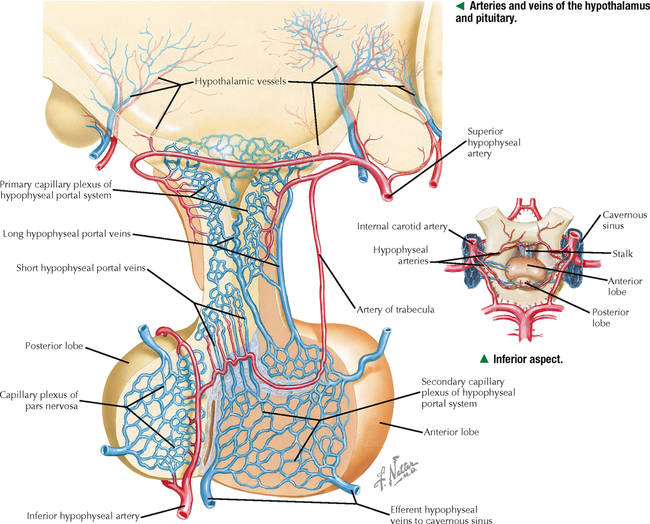
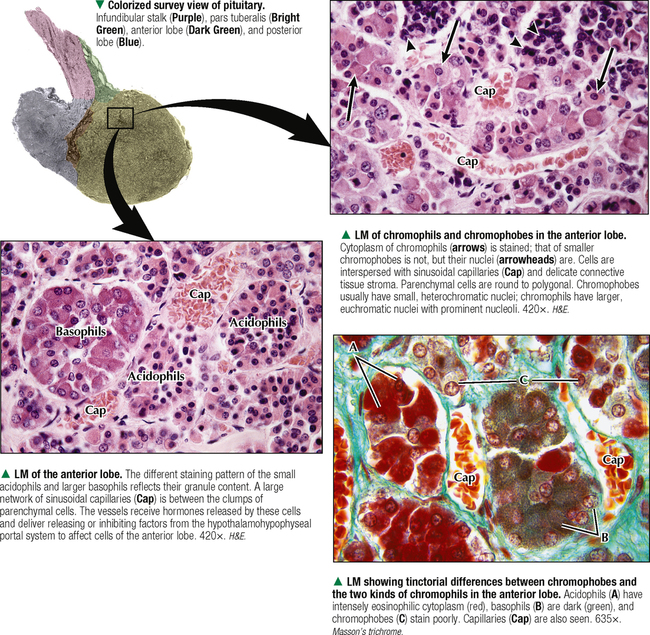
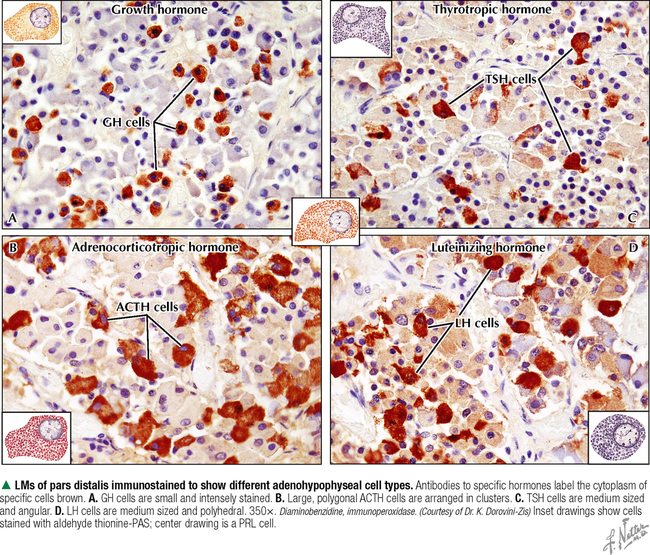
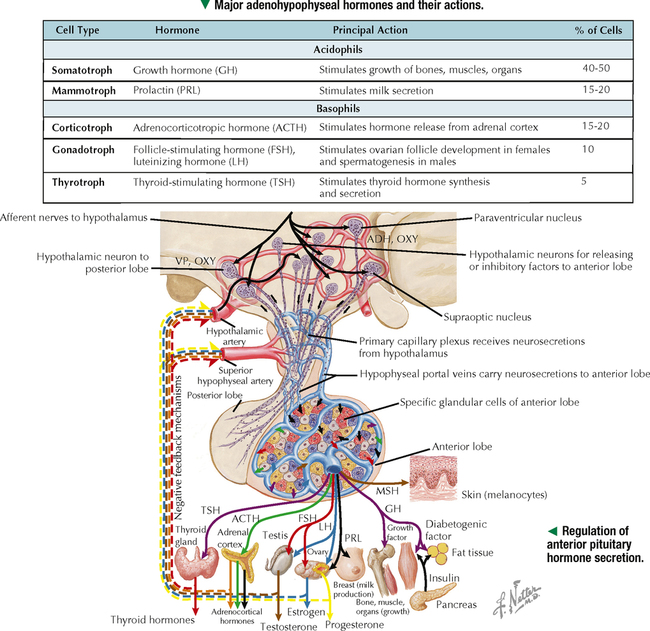
10 ENDOCRINE SYSTEM 10.1. Overview 10.2. Anatomy of the Pituitary 10.3. Development of the Pituitary 10.4. Divisions and Functions of the Pituitary 10.5. Blood Supply of the Pituitary 10.6. Histology and Function of the Pituitary Lobes 10.7. Histology of the Anterior Lobe: Chromophils and Chromophobes 10.8. Immunocytochemistry of Cells of the Pars Distalis 10.9. Functions of the Adenohypophysis 10.10. Ultrastructure of the Anterior Lobe 10.11. Functions of the Neurohypophysis 10.12. Histology of the Posterior Lobe 10.13. Ultrastructure and Function of the Posterior Lobe 10.14. Overview of the Thyroid and Parathyroid 10.15. Histology and Function of the Thyroid 10.16. Ultrastructure and Function of Thyroid Follicular Cells 10.17. Histology and Function of the Parathyroid 10.18. Histology and Ultrastructure of Parathyroid Chief Cells 10.19. Overview of the Adrenal and Its Blood Supply 10.20. Development of the Adrenal 10.21. Histology and Histochemistry of the Adrenal 10.22. Histology and Function of the Adrenal Cortex and Medulla 10.23. Ultrastructure of Spongiocytes in the Zona Fasciculata 10.24. Ultrastructure of Chromaffin Cells in the Adrenal Medulla 10.25. Overview and Histology of Islets of Langerhans 10.26. Immunocytochemistry of Islets of Langerhans 10.27. Ultrastructure of Islets of Langerhans 10.28. Ultrastructure and Function of Beta Cells 10.29. Histology of the Pineal 10.1 OVERVIEW The endocrine system comprises glands and tissues composed of parenchymal cells, which synthesize and secrete products called hormones. The term hormone derives from a Greek word meaning to set in motion. Like the nervous system, the endocrine system has diverse regulatory functions that control and coordinate activities of many other organs and tissues. Because the two systems have integrally linked functions, they are often termed the neuroendocrine system. The major endocrine glands—or ductless glands—are the pituitary (or hypophysis), hypothalamus, thyroid, parathyroids, adrenals, islets of Langerhans, and pineal. The placenta, which exists only during pregnancy, also elaborates several hormones, and other organs such as the heart, kidneys, thymus, gonads (testis and ovary), and intestines have isolated single cells or groups of cells with endocrine functions. Unlike exocrine glands that deliver secretions to a surface by ducts, endocrine glands lack ducts and release hormones into interstitial connective tissue. Hormones then pass to the blood or lymphatic circulation, their secretion usually being controlled by feedback mechanisms. Hormones are chemically diverse molecules—modified amino acids, peptides, glycoproteins, steroids, or biogenic amines—that typically affect target cells at sites distant from the site of release. Hormones usually bind target receptors either on cell membranes or within cells. Most endocrine glands consist of cords of parenchymal cells closely associated with a very rich vascular supply composed of a network of fenestrated capillaries and a relatively small amount of stroma. Hormone-secreting cells are usually, but not always, epithelial cells that abut walls of blood or lymphatic vessels. Nonepithelial cells that perform endocrine functions include atrial myocardial cells in the heart, smooth muscle cells in the juxtaglomerular apparatus of the kidney, and fat cells in adipose tissue. 10.2 ANATOMY OF THE PITUITARY The pituitary, or hypophysis, is often called the master endocrine gland because its hormones regulate physiologic activities of many other endocrine glands and tissues. It affects the growth, differentiation, and functions of many parts of the body, either directly by its own hormones or indirectly via secretions of other glands under its control. The size of a slightly flat grape or pea, it weighs 500–900 mg in adults; it is slightly larger in women, especially during pregnancy. The pituitary is strategically located at the base of the brain, at the hypothalamus, where the pituitary (or infundibular) stalk, to which it has extensive vascular and neural connections, suspends it. It lies in the midline in a depression of the sphenoid bone, the sella turcica, and is thus well protected. It has several important gross anatomic relationships. It is close anterolaterally to the optic chiasm and optic nerves, so pituitary lesions or tumors impinging on these structures can cause significant visual deficits. The sphenoid air sinus lies inferiorly to the gland, which allows relatively easy transsphenoidal surgical access to it. The superior aspect is covered by a thickened extension of the dura mater, named the diaphragmatic sellae, and is surrounded by a thin connective tissue capsule. It releases secretions into the bloodstream and is thus richly vascularized. The pituitary and the hypothalamus together make up a complex neuroendocrine circuit whose functions are closely linked. Despite its small size, the pituitary is essential to life. 10.3 DEVELOPMENT OF THE PITUITARY The pituitary has a dual embryonic origin, a combination of two distinct tissues. Early in gestation, the adenohypophysis—or anterior pituitary—arises as a dorsal outpocketing of thickened oral ectoderm called the Rathke pouch. By week 6 of gestation, it pinches off from the roof of the oral cavity and migrates to a site just anterior to a simultaneous downgrowth of neural ectoderm called the infundibular process. Cell proliferation in the anterior wall of the Rathke pouch then gives rise to the main part of the anterior lobe, which is the pars distalis. The terminal end of the ventral downgrowth of neural ectoderm becomes solid. Its cells give rise to the pars nervosa (posterior lobe) of the neurohypophysis (posterior pituitary), which keeps its neural connection to the brain. The two tissues become closely apposed but their microscopic structure remains different, reflecting the developmental dichotomy. Remnants of the Rathke pouch may persist as either a vestigial cleft or colloid-filled cysts at the anterior border with the neurohypophysis. The dorsal wall of the cleft fuses with the adjoining part of the posterior lobe to make up the small pars intermedia. In adults, the gland measures 1.2–1.5 cm in the transverse plane and about 1 cm in the sagittal plane. Its size varies greatly throughout life, however, depending on physiologic states. CLINICAL POINT About 10%-15% of intracranial tumors are usually benign pituitary adenomas of the adenohypophysis. Rarely malignant, they may be secretory or nonsecretory. They may arise from multiple oncogene abnormalities, such as G-protein and ras gene mutations, p53 gene deletions, and mutations that lead to multiple endocrine neoplasia. One type of secretory adenoma that usually requires surgery causes acromegaly in adults and gigantism in children. It is due to overproduction of growth hormone. Another type causes Cushing disease—an excess of corticotropin (ACTH)—which leads to overproduction of cortisol by adrenal glands. 10.4 DIVISIONS AND FUNCTIONS OF THE PITUITARY The adult adenohypophysis and neurohypophysis are each divided into three parts. The adenohypophysis, made of glandular epithelium, consists of the anterior lobe (pars distalis), the largest part; pars tuberalis, a thin collar of tissue surrounding the infundibular stalk; and pars intermedia (intermediate lobe), a narrow rudimentary band just posterior to the vestigial cleft and in contact with the posterior lobe. The neurohypophysis, made of neural tissue, comprises the posterior lobe, the main, most expanded part; median eminence, the upper part that attaches the gland to the hypothalamus; and connecting infundibular stalk. The median eminence is partly encircled by the pars tuberalis, which links it via a network of many capillaries to the anterior lobe. The adenohypophysis synthesizes and secretes several polypeptide and glycoprotein hormones; the neurohypophysis (by way of modified neurons from the hypothalamus) secretes two peptide hormones. All hormones enter the systemic circulation and are taken to distant target tissues to regulate functions. CLINICAL POINT Acromegaly in adults—a potentially life-threatening metabolic disorder usually caused by increased and unregulated growth hormone (GH) production due to a GH-secreting (somatotroph) adenoma—is named from Greek (acro: extremities; megaly: enlargement) because of its distinctive features: abnormal growth of hands and feet. Thickening of cranial bones, mandible, soft tissues, and internal organs (visceromegaly) are other symptoms. Such clinical signs are caused by chronic elevated serum levels of GH that stimulate hepatic secretion of insulin-like growth factor (IGF-1). Left untreated, serious complications and premature death occur, mainly from cardiovascular disease. To improve the prognosis and quality of life, treatment is surgery to excise the tumor and sometimes follow-up radiation and pharmacologic therapy (e.g., long-acting somatostatin analogs, GH receptor agonists) to reduce growth-promoting effects of GH. 10.5 BLOOD SUPPLY OF THE PITUITARY Two paired hypophyseal arteries—branches of internal carotids—have profuse anastomoses on the gland’s surface. The superior hypophyseal arteries, from above, bring blood to the anterior lobe by first forming a primary capillary plexus made of vascular loops in the area of the median eminence and pars tuberalis. These vessels give rise to a network of portal venules—the hypophyseal portal system—which crosses the ventral aspect of the pituitary stalk to drain into a secondary plexus of sinusoidal fenestrated capillaries in the anterior lobe. This portal system is critical for control of the adenohypophysis by neurosecretions from hypothalamic neurons that convey releasing and inhibiting hormones to the primary plexus. Neurosecretions reach the secondary plexus to regulate release of specific adenohypophysis hormones, which are also secreted into the secondary capillary plexus. Small efferent veins, in turn, drain into cavernous sinuses surrounding the gland. Inferior hypophyseal arteries, below, carry arterial blood to the posterior lobe. They drain into a plexus of sinusoidal fenestrated capillaries that take blood via efferent hypophyseal veins to the cavernous sinus. An important branch of the superior hypophyseal artery, the artery of the trabecula, bypasses the portal circulation and forms small capillary loops in the pars intermedia, which anastomose with capillaries in the anterior lobe. The anterior and posterior lobes are richly vascularized, but the pars intermedia is not. CLINICAL POINT Sheehan syndrome (postpartum hypopituitarism) affects women with life-threatening blood loss during childbirth. Because of ischemic necrosis of the anterior pituitary caused by severe postpartum hemorrhage, symptoms include cessation of lactation (agalactorrhea) and menstrual periods (amenorrhea, hypotension, and fatigue). Because of advances in obstetrics in industrialized countries, it is a rare complication, but in other parts of the world, it is a major threat to pregnant women and a common cause of hypopituitarism. In pregnancy, the anterior pituitary nearly doubles in size but without simultaneous increase in blood supply, so the gland becomes vulnerable to anoxia and infarction. If profound postpartum bleeding occurs, blood supply to the anterior lobe is inadequate, resulting in parenchymal cell necrosis. Lifetime hormone replacement therapy (e.g., estrogen, corticosteroids, thyroid hormones) usually improves prognosis. 10.6 HISTOLOGY AND FUNCTION OF THE PITUITARY LOBES Low magnification resolves the contrasting histologic structure between the pars distalis of the anterior lobe and pars nervosa of the posterior lobe. The anterior lobe is glandular epithelium, which stains dark because of its many, tightly packed nucleated parenchymal cells. The posterior lobe is more lightly stained because it is typically made of nervous tissue. In the intermediate lobe, at the border with the posterior lobe, rudiments of the Rathke pouch persist as accumulations of small colloid-filled cysts. Showing great size variation among species, the intermediate lobe constitutes less than 2% of the adult human pituitary. This lobe is rudimentary in humans and its function in adults is uncertain, but it consists of either isolated groups of low columnar epithelial cells or a discontinuous epithelial layer, which often surrounds colloid-filled follicles, and contains basophilic parenchymal cells and a few scattered, lightly stained polygonal cells. Cells in this lobe produce melanocyte-stimulating hormone and the opiate peptide β-endorphin. Basophilic cells from the intermediate lobe often invade the posterior lobe. 10.7 HISTOLOGY OF THE ANTERIOR LOBE: CHROMOPHILS AND CHROMOPHOBES About 75% of the adenohypophysis is anterior lobe. It consists of clumps or cords of glandular epithelial cells in close relation to a network of sinusoidal capillaries with large and irregular lumina. Scant loose connective tissue is made of delicate reticular fiber stroma, which supports glandular cells and sinusoid walls. Hematoxylin and eosin (H&E) reveals two distinct parenchymal cell types: chromophils (large, have secretory granules, stain intensely) and chromophobes (smaller, have few or no secretory granules, stain faintly). Chromophobes have less cytoplasm than do chromophils and may be quiescent, degranulated, or undifferentiated cells. Chromophils can be distinguished as acidophils or basophils on the basis of their cytoplasmic affinity for acid or basic dyes and on the tinctorial properties of their secretory granules. Acidophils, typically smaller cells with smoothly refractive cytoplasm, secrete two polypeptide hormones. The larger basophils are more granular and secrete four major polypeptide hormones. Via routine stains, proportions of glandular cell types are about 40% acidophils, 10% basophils, and 50% chromophobes. Their distribution differs regionally and varies locally. Immunocytochemistry with specific antibodies has allowed more precise identification of these cells and their hormone content. A functional nomenclature is now routinely used to designate cell types according to the secreted hormone or target organ. 10.8 IMMUNOCYTOCHEMISTRY OF CELLS OF THE PARS DISTALIS Immunocytochemistry uses antibodies to protein secretory products and allows identification of five adenohypophyseal cell types on the basis of intracellular localization of one or more hormones. Use of immunocytochemistry helps clarify the normal regional distribution of cells, correlates structure to function, and aids tumor diagnosis. Small, ovoid somatotrophs (GH [growth hormone] cells) are mostly in lateral wings of the anterior lobe. Their abundant secretory granules produce intense immunostaining. Corticotrophs (ACTH [adrenocorticotropic hormone, or corticotropin] cells) are most numerous in middle and posterior parts of the gland, typically in clusters. These polygonal, medium to large cells stain for corticotropin, melanocyte-stimulating hormone, endorphin, and enkephalin. Many cells have an unstained area near the nucleus, which indicates a large lysosome. Medium-sized, angular thyrotrophs (TSH [thyroid-stimulating hormone, or thyrotropin] cells) are found in small groups in an anterior wedge part of the gland. Gonadotrophs (FSH [follicle-stimulating hormone] and LH [luteinizing hormone] cells) are evenly distributed throughout the anterior lobe. The same or different cells may produce the two hormones. Large, polyhedral mammotrophs (known as either PRL [prolactin] or LTH [lactogenic hormone] cells) are in posterior parts of the lateral wings. They are densely or sparsely granulated. In pregnant or lactating women, PRL cells usually undergo significant hyperplasia and hypertrophy. CLINICAL POINT Immunocytochemistry helps detect pituitary adenomas and allows diagnosis on the basis of hormones produced by neoplastic cells. Prolactinoma, accounting for 30% of all neoplastic pituitary tumors, is the most common type. A tumor of mammotrophs, it leads to amenorrhea, infertility, osteopenia, and galactorrhea in women and erectile dysfunction and loss of libido in men. Amyloid deposits and calcified spherites (or psammoma bodies) accompany excessive synthesis and secretion of prolactin. Treatment with the dopamine agonist bromocriptine reduces tumor size and inhibits prolactin secretion. Tumors larger than 10 mm in diameter (macroadenomas) require surgery or radiation. Only gold members can continue reading. Log In or Register to continue Share this: Share on X (Opens in new window) X Share on Facebook (Opens in new window) Facebook Like this:Like Loading… Related Related posts: CARDIOVASCULAR SYSTEM RESPIRATORY SYSTEM SPECIAL SENSES LOWER DIGESTIVE SYSTEM Stay updated, free articles. Join our Telegram channel Join Tags: Netters Essential Histology Jun 18, 2016 | Posted by admin in HISTOLOGY | Comments Off on ENDOCRINE SYSTEM Full access? Get Clinical Tree
10 ENDOCRINE SYSTEM 10.1. Overview 10.2. Anatomy of the Pituitary 10.3. Development of the Pituitary 10.4. Divisions and Functions of the Pituitary 10.5. Blood Supply of the Pituitary 10.6. Histology and Function of the Pituitary Lobes 10.7. Histology of the Anterior Lobe: Chromophils and Chromophobes 10.8. Immunocytochemistry of Cells of the Pars Distalis 10.9. Functions of the Adenohypophysis 10.10. Ultrastructure of the Anterior Lobe 10.11. Functions of the Neurohypophysis 10.12. Histology of the Posterior Lobe 10.13. Ultrastructure and Function of the Posterior Lobe 10.14. Overview of the Thyroid and Parathyroid 10.15. Histology and Function of the Thyroid 10.16. Ultrastructure and Function of Thyroid Follicular Cells 10.17. Histology and Function of the Parathyroid 10.18. Histology and Ultrastructure of Parathyroid Chief Cells 10.19. Overview of the Adrenal and Its Blood Supply 10.20. Development of the Adrenal 10.21. Histology and Histochemistry of the Adrenal 10.22. Histology and Function of the Adrenal Cortex and Medulla 10.23. Ultrastructure of Spongiocytes in the Zona Fasciculata 10.24. Ultrastructure of Chromaffin Cells in the Adrenal Medulla 10.25. Overview and Histology of Islets of Langerhans 10.26. Immunocytochemistry of Islets of Langerhans 10.27. Ultrastructure of Islets of Langerhans 10.28. Ultrastructure and Function of Beta Cells 10.29. Histology of the Pineal 10.1 OVERVIEW The endocrine system comprises glands and tissues composed of parenchymal cells, which synthesize and secrete products called hormones. The term hormone derives from a Greek word meaning to set in motion. Like the nervous system, the endocrine system has diverse regulatory functions that control and coordinate activities of many other organs and tissues. Because the two systems have integrally linked functions, they are often termed the neuroendocrine system. The major endocrine glands—or ductless glands—are the pituitary (or hypophysis), hypothalamus, thyroid, parathyroids, adrenals, islets of Langerhans, and pineal. The placenta, which exists only during pregnancy, also elaborates several hormones, and other organs such as the heart, kidneys, thymus, gonads (testis and ovary), and intestines have isolated single cells or groups of cells with endocrine functions. Unlike exocrine glands that deliver secretions to a surface by ducts, endocrine glands lack ducts and release hormones into interstitial connective tissue. Hormones then pass to the blood or lymphatic circulation, their secretion usually being controlled by feedback mechanisms. Hormones are chemically diverse molecules—modified amino acids, peptides, glycoproteins, steroids, or biogenic amines—that typically affect target cells at sites distant from the site of release. Hormones usually bind target receptors either on cell membranes or within cells. Most endocrine glands consist of cords of parenchymal cells closely associated with a very rich vascular supply composed of a network of fenestrated capillaries and a relatively small amount of stroma. Hormone-secreting cells are usually, but not always, epithelial cells that abut walls of blood or lymphatic vessels. Nonepithelial cells that perform endocrine functions include atrial myocardial cells in the heart, smooth muscle cells in the juxtaglomerular apparatus of the kidney, and fat cells in adipose tissue. 10.2 ANATOMY OF THE PITUITARY The pituitary, or hypophysis, is often called the master endocrine gland because its hormones regulate physiologic activities of many other endocrine glands and tissues. It affects the growth, differentiation, and functions of many parts of the body, either directly by its own hormones or indirectly via secretions of other glands under its control. The size of a slightly flat grape or pea, it weighs 500–900 mg in adults; it is slightly larger in women, especially during pregnancy. The pituitary is strategically located at the base of the brain, at the hypothalamus, where the pituitary (or infundibular) stalk, to which it has extensive vascular and neural connections, suspends it. It lies in the midline in a depression of the sphenoid bone, the sella turcica, and is thus well protected. It has several important gross anatomic relationships. It is close anterolaterally to the optic chiasm and optic nerves, so pituitary lesions or tumors impinging on these structures can cause significant visual deficits. The sphenoid air sinus lies inferiorly to the gland, which allows relatively easy transsphenoidal surgical access to it. The superior aspect is covered by a thickened extension of the dura mater, named the diaphragmatic sellae, and is surrounded by a thin connective tissue capsule. It releases secretions into the bloodstream and is thus richly vascularized. The pituitary and the hypothalamus together make up a complex neuroendocrine circuit whose functions are closely linked. Despite its small size, the pituitary is essential to life. 10.3 DEVELOPMENT OF THE PITUITARY The pituitary has a dual embryonic origin, a combination of two distinct tissues. Early in gestation, the adenohypophysis—or anterior pituitary—arises as a dorsal outpocketing of thickened oral ectoderm called the Rathke pouch. By week 6 of gestation, it pinches off from the roof of the oral cavity and migrates to a site just anterior to a simultaneous downgrowth of neural ectoderm called the infundibular process. Cell proliferation in the anterior wall of the Rathke pouch then gives rise to the main part of the anterior lobe, which is the pars distalis. The terminal end of the ventral downgrowth of neural ectoderm becomes solid. Its cells give rise to the pars nervosa (posterior lobe) of the neurohypophysis (posterior pituitary), which keeps its neural connection to the brain. The two tissues become closely apposed but their microscopic structure remains different, reflecting the developmental dichotomy. Remnants of the Rathke pouch may persist as either a vestigial cleft or colloid-filled cysts at the anterior border with the neurohypophysis. The dorsal wall of the cleft fuses with the adjoining part of the posterior lobe to make up the small pars intermedia. In adults, the gland measures 1.2–1.5 cm in the transverse plane and about 1 cm in the sagittal plane. Its size varies greatly throughout life, however, depending on physiologic states. CLINICAL POINT About 10%-15% of intracranial tumors are usually benign pituitary adenomas of the adenohypophysis. Rarely malignant, they may be secretory or nonsecretory. They may arise from multiple oncogene abnormalities, such as G-protein and ras gene mutations, p53 gene deletions, and mutations that lead to multiple endocrine neoplasia. One type of secretory adenoma that usually requires surgery causes acromegaly in adults and gigantism in children. It is due to overproduction of growth hormone. Another type causes Cushing disease—an excess of corticotropin (ACTH)—which leads to overproduction of cortisol by adrenal glands. 10.4 DIVISIONS AND FUNCTIONS OF THE PITUITARY The adult adenohypophysis and neurohypophysis are each divided into three parts. The adenohypophysis, made of glandular epithelium, consists of the anterior lobe (pars distalis), the largest part; pars tuberalis, a thin collar of tissue surrounding the infundibular stalk; and pars intermedia (intermediate lobe), a narrow rudimentary band just posterior to the vestigial cleft and in contact with the posterior lobe. The neurohypophysis, made of neural tissue, comprises the posterior lobe, the main, most expanded part; median eminence, the upper part that attaches the gland to the hypothalamus; and connecting infundibular stalk. The median eminence is partly encircled by the pars tuberalis, which links it via a network of many capillaries to the anterior lobe. The adenohypophysis synthesizes and secretes several polypeptide and glycoprotein hormones; the neurohypophysis (by way of modified neurons from the hypothalamus) secretes two peptide hormones. All hormones enter the systemic circulation and are taken to distant target tissues to regulate functions. CLINICAL POINT Acromegaly in adults—a potentially life-threatening metabolic disorder usually caused by increased and unregulated growth hormone (GH) production due to a GH-secreting (somatotroph) adenoma—is named from Greek (acro: extremities; megaly: enlargement) because of its distinctive features: abnormal growth of hands and feet. Thickening of cranial bones, mandible, soft tissues, and internal organs (visceromegaly) are other symptoms. Such clinical signs are caused by chronic elevated serum levels of GH that stimulate hepatic secretion of insulin-like growth factor (IGF-1). Left untreated, serious complications and premature death occur, mainly from cardiovascular disease. To improve the prognosis and quality of life, treatment is surgery to excise the tumor and sometimes follow-up radiation and pharmacologic therapy (e.g., long-acting somatostatin analogs, GH receptor agonists) to reduce growth-promoting effects of GH. 10.5 BLOOD SUPPLY OF THE PITUITARY Two paired hypophyseal arteries—branches of internal carotids—have profuse anastomoses on the gland’s surface. The superior hypophyseal arteries, from above, bring blood to the anterior lobe by first forming a primary capillary plexus made of vascular loops in the area of the median eminence and pars tuberalis. These vessels give rise to a network of portal venules—the hypophyseal portal system—which crosses the ventral aspect of the pituitary stalk to drain into a secondary plexus of sinusoidal fenestrated capillaries in the anterior lobe. This portal system is critical for control of the adenohypophysis by neurosecretions from hypothalamic neurons that convey releasing and inhibiting hormones to the primary plexus. Neurosecretions reach the secondary plexus to regulate release of specific adenohypophysis hormones, which are also secreted into the secondary capillary plexus. Small efferent veins, in turn, drain into cavernous sinuses surrounding the gland. Inferior hypophyseal arteries, below, carry arterial blood to the posterior lobe. They drain into a plexus of sinusoidal fenestrated capillaries that take blood via efferent hypophyseal veins to the cavernous sinus. An important branch of the superior hypophyseal artery, the artery of the trabecula, bypasses the portal circulation and forms small capillary loops in the pars intermedia, which anastomose with capillaries in the anterior lobe. The anterior and posterior lobes are richly vascularized, but the pars intermedia is not. CLINICAL POINT Sheehan syndrome (postpartum hypopituitarism) affects women with life-threatening blood loss during childbirth. Because of ischemic necrosis of the anterior pituitary caused by severe postpartum hemorrhage, symptoms include cessation of lactation (agalactorrhea) and menstrual periods (amenorrhea, hypotension, and fatigue). Because of advances in obstetrics in industrialized countries, it is a rare complication, but in other parts of the world, it is a major threat to pregnant women and a common cause of hypopituitarism. In pregnancy, the anterior pituitary nearly doubles in size but without simultaneous increase in blood supply, so the gland becomes vulnerable to anoxia and infarction. If profound postpartum bleeding occurs, blood supply to the anterior lobe is inadequate, resulting in parenchymal cell necrosis. Lifetime hormone replacement therapy (e.g., estrogen, corticosteroids, thyroid hormones) usually improves prognosis. 10.6 HISTOLOGY AND FUNCTION OF THE PITUITARY LOBES Low magnification resolves the contrasting histologic structure between the pars distalis of the anterior lobe and pars nervosa of the posterior lobe. The anterior lobe is glandular epithelium, which stains dark because of its many, tightly packed nucleated parenchymal cells. The posterior lobe is more lightly stained because it is typically made of nervous tissue. In the intermediate lobe, at the border with the posterior lobe, rudiments of the Rathke pouch persist as accumulations of small colloid-filled cysts. Showing great size variation among species, the intermediate lobe constitutes less than 2% of the adult human pituitary. This lobe is rudimentary in humans and its function in adults is uncertain, but it consists of either isolated groups of low columnar epithelial cells or a discontinuous epithelial layer, which often surrounds colloid-filled follicles, and contains basophilic parenchymal cells and a few scattered, lightly stained polygonal cells. Cells in this lobe produce melanocyte-stimulating hormone and the opiate peptide β-endorphin. Basophilic cells from the intermediate lobe often invade the posterior lobe. 10.7 HISTOLOGY OF THE ANTERIOR LOBE: CHROMOPHILS AND CHROMOPHOBES About 75% of the adenohypophysis is anterior lobe. It consists of clumps or cords of glandular epithelial cells in close relation to a network of sinusoidal capillaries with large and irregular lumina. Scant loose connective tissue is made of delicate reticular fiber stroma, which supports glandular cells and sinusoid walls. Hematoxylin and eosin (H&E) reveals two distinct parenchymal cell types: chromophils (large, have secretory granules, stain intensely) and chromophobes (smaller, have few or no secretory granules, stain faintly). Chromophobes have less cytoplasm than do chromophils and may be quiescent, degranulated, or undifferentiated cells. Chromophils can be distinguished as acidophils or basophils on the basis of their cytoplasmic affinity for acid or basic dyes and on the tinctorial properties of their secretory granules. Acidophils, typically smaller cells with smoothly refractive cytoplasm, secrete two polypeptide hormones. The larger basophils are more granular and secrete four major polypeptide hormones. Via routine stains, proportions of glandular cell types are about 40% acidophils, 10% basophils, and 50% chromophobes. Their distribution differs regionally and varies locally. Immunocytochemistry with specific antibodies has allowed more precise identification of these cells and their hormone content. A functional nomenclature is now routinely used to designate cell types according to the secreted hormone or target organ. 10.8 IMMUNOCYTOCHEMISTRY OF CELLS OF THE PARS DISTALIS Immunocytochemistry uses antibodies to protein secretory products and allows identification of five adenohypophyseal cell types on the basis of intracellular localization of one or more hormones. Use of immunocytochemistry helps clarify the normal regional distribution of cells, correlates structure to function, and aids tumor diagnosis. Small, ovoid somatotrophs (GH [growth hormone] cells) are mostly in lateral wings of the anterior lobe. Their abundant secretory granules produce intense immunostaining. Corticotrophs (ACTH [adrenocorticotropic hormone, or corticotropin] cells) are most numerous in middle and posterior parts of the gland, typically in clusters. These polygonal, medium to large cells stain for corticotropin, melanocyte-stimulating hormone, endorphin, and enkephalin. Many cells have an unstained area near the nucleus, which indicates a large lysosome. Medium-sized, angular thyrotrophs (TSH [thyroid-stimulating hormone, or thyrotropin] cells) are found in small groups in an anterior wedge part of the gland. Gonadotrophs (FSH [follicle-stimulating hormone] and LH [luteinizing hormone] cells) are evenly distributed throughout the anterior lobe. The same or different cells may produce the two hormones. Large, polyhedral mammotrophs (known as either PRL [prolactin] or LTH [lactogenic hormone] cells) are in posterior parts of the lateral wings. They are densely or sparsely granulated. In pregnant or lactating women, PRL cells usually undergo significant hyperplasia and hypertrophy. CLINICAL POINT Immunocytochemistry helps detect pituitary adenomas and allows diagnosis on the basis of hormones produced by neoplastic cells. Prolactinoma, accounting for 30% of all neoplastic pituitary tumors, is the most common type. A tumor of mammotrophs, it leads to amenorrhea, infertility, osteopenia, and galactorrhea in women and erectile dysfunction and loss of libido in men. Amyloid deposits and calcified spherites (or psammoma bodies) accompany excessive synthesis and secretion of prolactin. Treatment with the dopamine agonist bromocriptine reduces tumor size and inhibits prolactin secretion. Tumors larger than 10 mm in diameter (macroadenomas) require surgery or radiation. Only gold members can continue reading. Log In or Register to continue Share this: Share on X (Opens in new window) X Share on Facebook (Opens in new window) Facebook Like this:Like Loading… Related Related posts: CARDIOVASCULAR SYSTEM RESPIRATORY SYSTEM SPECIAL SENSES LOWER DIGESTIVE SYSTEM Stay updated, free articles. Join our Telegram channel Join