2 The development of a single fertilised egg cell to form a complex multicellular organism involves cellular replication, growth and progressive specialisation (differentiation) for a variety of functions. The fertilised egg (zygote) divides by a process known as mitosis to produce two genetically identical daughter cells, each of which divides to produce two more daughter cells and so on. Some of these daughter cells progressively specialise and eventually produce the terminally differentiated cells of mature tissues, such as muscle or skin cells. Most tissues, however, retain a population of relatively undifferentiated cells (stem cells) that are able to divide and replace the differentiated cell population as required. The interval between mitotic divisions is known as the cell cycle. All body cells divide by mitosis except for male and female germ cells, which divide by meiosis to produce gametes (see Fig. 2.6). Cell division and differentiation are balanced by cell death, both during the development and growth of the immature organism and in the mature adult. In these circumstances, cell death occurs by a mechanism known as apoptosis (see Fig. 2.7). FIG. 2.1 The cell cycle FIG. 2.2 Chromosomes during mitosis (illustrations opposite) FIG. 2.3 Mitosis (illustration (b) opposite) • Prometaphase. Dissolution of the nuclear envelope marks the beginning of prometaphase. The mitotic spindle then moves into the nuclear area and each duplicated chromosome becomes attached at a site called the kinetochore to another group of microtubules of the mitotic spindle (kinetochore or chromosome microtubules). The kinetochore is a DNA and protein structure on each duplicated chromosome, located at the centromere, the structure which binds the duplicated chromosomes (chromatids) together (see also diagram (d), Fig. 2.2). Other microtubules attach the chromosome arms to the spindle.
Cell cycle and replication
Introduction
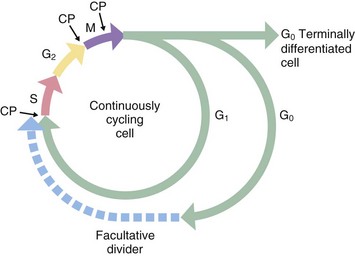
Historically, only two phases of the cell cycle were recognised: a relatively short mitotic phase (M phase) and a non-dividing phase (interphase), which usually occupies most of the life cycle of the cell. However, there is a discrete period during interphase when nuclear DNA is replicated; this phase, described as the synthesis or S phase, is completed some time before the onset of mitosis. Thus interphase may be divided into three separate phases. Between the end of the M phase and the beginning of the S phase is the first gap or G1 phase; this is usually much longer than the other phases of the cell cycle. During the G1 phase, cells differentiate and perform their specialised functions as part of the whole tissue. The interval between the end of the S phase and the beginning of the M phase, the second gap or G2 phase, is relatively short and is the period in which cells prepare for mitotic division.
Stem cells in some tissues progress continually through the cell cycle to accommodate tissue growth or cell turnover, whereas terminally differentiated cells leave the cell cycle after the M phase and enter a state of continuous differentiated function designated as G0 phase. Facultative dividers enter the G0 phase but retain the capacity to re-enter the cell cycle when suitably stimulated. Liver cells are a prominent example of facultative dividers with differentiated hepatocytes acting as stem cells in cases of massive liver injury. Some liver cells appear to enter a protracted G2 phase in which they perform their normal differentiated functions despite the presence of a duplicated complement of DNA. These cells may be seen histologically as binucleate cells.
In general, the S, G and M phases of the cell cycle are relatively constant in duration, each taking up to several hours to complete, whereas the G1 phase is highly variable, in some cases lasting for several days or weeks. M phase typically lasts about 1 hour, while S phase takes 10 to 12 hours to complete. The G0 phase may last for the entire lifespan of the organism.
During mitosis, various checkpoints CP prevent progression to the next phase before the previous one is completed. For example, the metaphase checkpoint prevents progression to anaphase before all the chromosomes are properly connected to the mitotic spindle and lined up at the cell equator; this prevents unequal distribution of the chromosomes to the two daughter cells.
Mitosis
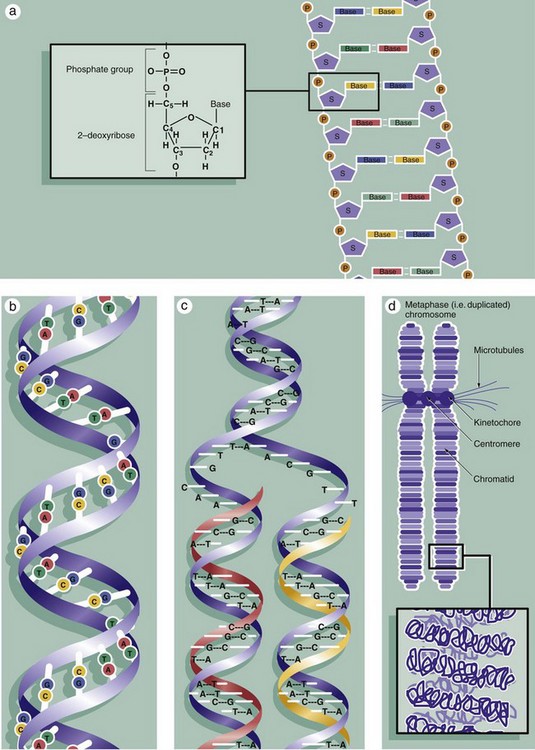
The nuclei of all somatic cells of an individual contain the same fixed complement of deoxyribonucleic acid (DNA), a quantity called the genome. The DNA is arranged into chromosomes, with each species having a set number. DNA is a very large molecular weight polymer consisting of many deoxyribonucleotides with a double-stranded structure. Each strand consists of a backbone of alternating deoxyribose S and phosphate P moieties. Each deoxyribose unit is covalently bound to a purine or pyrimidine base, which is in turn non-covalently linked to a complementary base on the other strand, thus linking the strands together. The bases are of four types, adenine A, cytosine C, thymine T and guanine G, with adenine only linking to thymine and cytosine only linking to guanine, thus making each strand complementary to the other, diagram (a). Linked in this way, the strands assume a double helical conformation around a common axis, the internucleotide phosphodiester bonds running in opposite directions (i.e. antiparallel) and the planes of the linked bases lying at right angles to the axis, diagram (b). The sequence of bases in either strand of the DNA molecule forms the genetic code for the individual. The bases are read in groups of three called codons, each coding for one amino acid. In human cells, there are 46 chromosomes (the diploid number) comprising 22 homologous pairs, the autosomes, and 2 sex chromosomes, either XX in the female or XY in the male. The members of each pair of autosomes have the same length of DNA and code for the same proteins.
Histologically, individual chromosomes are not visible within the cell nucleus during interphase. During S phase, each chromosome is duplicated, as shown in diagram (c). The resulting identical chromosomes, now known as sister chromatids, remain attached to one another at a point called the centromere and become even more tightly coiled and condensed when they may be visualised with the light microscope (see Figs 2.3 and 2.4). Accurate reproduction of the DNA strand is vital and multiple complex mechanisms exist to prevent mistakes (mutations) from occurring.
The extremely long DNA molecule making up each chromosome binds to a range of histone proteins that hold the chromosome in a supercoiled and folded conformation, compact enough to be accommodated within the nucleus (not illustrated). Thus the 2 nm–diameter double helix is coiled and packed through several orders of three-dimensional complexity to form an elongated structure some 300 nm in diameter and very much shorter in length than the otherwise uncoiled molecule would be. This is the form in which the chromosome is structured during the G1 and G0 phases of the cell cycle, during which gene transcription (the prerequisite for protein synthesis) occurs. Histone proteins are also reduplicated during S phase so that the sister chromatids will have their own complement of histones.
Diagram (d) shows further detail of the structure of mitotic chromosomes and their supercoiled three-dimensional structure. Note the position of the kinetochore (see Fig. 2.3) that provides attachment for the microtubules of the cell spindle during cell division and seems also to control the progression of mitosis. Examination of the chromosomes of dividing cells, karyotyping, can give diagnostic information about the chromosomal complement of an individual or of a malignant tumour (see Fig. 2.5).
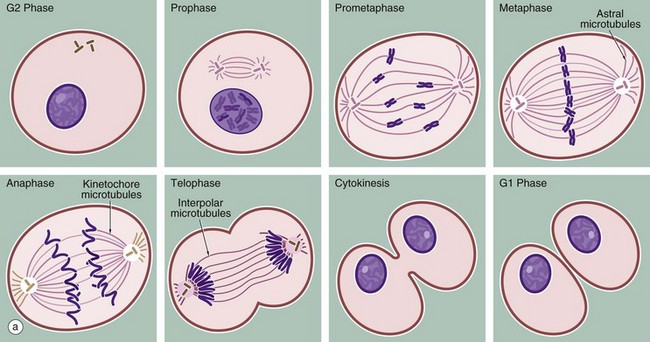
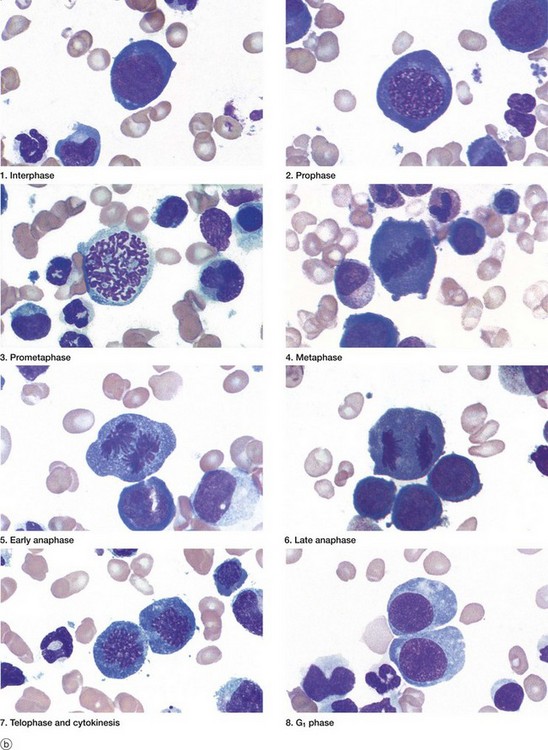
(a) Schematic diagram (b) Mitotic series Giemsa (HP)
The series of micrographs shown opposite illustrates the mitotic process in actively dividing immature blood cells from a smear preparation of human bone marrow. Mitosis is a continuous process that is traditionally divided into five phases, prophase, prometaphase, metaphase, anaphase and telophase, each stage being readily recognisable with the light microscope. Cell division requires the presence of a structure called the mitotic apparatus, which comprises a spindle of longitudinally arranged microtubules extending between a pair of centrioles (see Figs 1.17 and 1.18) at each pole of the dividing cell. The mitotic apparatus is visible within the cytoplasm only during M phase as it disaggregates shortly after completion of mitosis.
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Cell cycle and replication
