1Abbreviations: ARE, antioxidant-response element; BCO1, β-carotene-15,15′-oxygenase; BCO2, β-carotene-9′,10′-oxygenase; Cx43, connexin 43; GJC, gap junction communication; HDL, highdensity lipoprotein; HPLC, high-performance liquid chromatography; IGF, insulinlike growth factor; IGFBP, insulinlike growth factorbinding protein; ISX, intestine-specific homeobox; LDL, low-density lipoprotein; NHANES, National Health and Nutrition Examination Survey; NMR, nuclear magnetic resonance; Nrf2, nuclear factor E2-related factor 2; PPAR, peroxisome proliferator-activated receptor; PPRE, peroxisome proliferator-response element; RAR, retinoic acid receptor; RXR, retinoid X receptor; SNP, single nucleotide polymorphism; SR-B1, scavenger receptor class B type 1 protein transporter; UV, ultraviolet; VLDL, low-density lipoprotein.
HISTORICAL OVERVIEW
Carotenoids are lipophilic pigments that occur widely in plants, insects, fish, birds, algae, yeasts, and bacteria, and they carry out various important biologic functions. Studies on carotenoids began in 1831, when Wackenroder first isolated the crystalline yellow pigment carotene from carrots (Daucus carota), and studies continued in 1837, when Berzelius named the yellow pigments of autumn leaves xanthophylls (1). In the early twentieth century, the technique of chromatography was discovered, and carotenoid analysis underwent great development. Researchers discovered a large family of carotenoids and found that these substances were isoprenoid derivatives. In 1913, very close to the discovery of vitamin A by McCollum and Davis (see the chapter on vitamin A), Osborne and Mendel observed that the green parts of plants contained a relatively high amount of “fat-soluble A” activity. During the subsequent decade, Steenbock, Moore, and other investigators provided further information on the relationship between the fat-soluble A and yellow pigments from comparative studies of their growth-promoting activities in vivo.
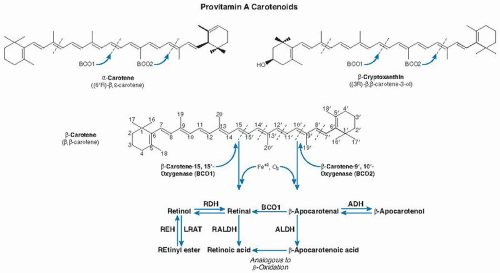
In 1930 and 1931, Karrer et al defined the chemical structures of both β-carotene and retinol purified from shark liver oil. These researchers determined that half of the chemical structure of β-carotene resembles the retinol structure. This finding led Karrer to suggest that the simple addition of two molecules of water at the central double bond of the carotene molecule should yield two molecules of retinol. However, it was not until 1965 that Goodman and Huang (2) and Olson and Hayaishi (3) independently demonstrated the enzymatic synthesis of retinal from β-carotene in cell-free extracts of rat liver and intestine. These investigators did not detect any other cleavage products and therefore suggested that a process took place by means of cleavage symmetrically at the β-carotene central double bond that required molecular oxygen. These investigators named the enzyme β-carotene-15,15′-oxygenase (BCO1).
It was another 35 years until Wyss et al (4) and von Lintig and Vogt (5), in different species, were able to achieve the molecular cloning of the BCO1 gene; these investigators subsequently provided a biochemical and structural characterization (6, 7, 8, 9, 10, 11). These studies firmly established that the central cleavage of provitamin A carotenoids is the main pathway leading to the formation of vitamin A (Fig. 31.1).
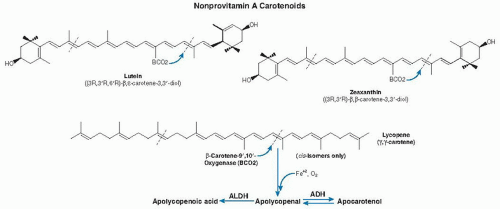
In 1954, Glover et al (12) proposed that β-carotene can undergo both central and eccentric cleavage. Eccentric cleavage is the asymmetric cleavage of carotenoids at positions outside the central double bond (see Fig. 31.1). The existence of the eccentric cleavage pathway for β-carotene had been controversial among scientists since the 1970s (13, 14, 15, 16), however, and it was not confirmed until the molecular identification of β-carotene-9′,10′-oxygenase (BCO2) in humans and mice by Kiefer et al (17). Investigators further showed that BCO2 preferentially cleaves nonprovitamin A carotenoids (cis-lycopene, lutein, and zeaxanthin) (18, 19) (Fig. 31.2).
Fig. 31.1. Metabolic pathway and chemical structures of the major provitamin A carotenoids (β-carotene, α-carotene, and β-cryptoxanthin) found in human plasma and tissues. Provitamin A carotenoids are symmetrically cleaved at the 15,15′ double bond by β-carotene-15,15′-oxygenase (BCO1), which produces one or two molecules of all-trans-retinal that can be oxidized to retinoic acid or reduced to retinol. Retinol can be converted to retinyl esters for storage. Provitamin A carotenoids can also be cleaved by β-carotene-9′,10′-oxygenase (BCO2) at either the 9,10 or 9′,10′ double bond, thus giving rise to β-apo-10′-carotenal and β-ionone. Apo-β-carotenals either can be precursors of vitamin A by further BCO2 cleavage or can be oxidized to their corresponding apo-β-carotenoic acids, which may then undergo a process similar to β-oxidation of fatty acids, to produce retinoic acid. ADH, alcohol dehydrogenase; ALDH, aldehyde dehydrogenase; LRAT, lecithin:retinol acyltransferase; RALDH, retinal dehydrogenase; RDH, retinol dehydrogenase; REH, retinyl ester hydrolase.
Data indicate that carotenoid metabolites, in addition to vitamin A, may participate in specific biologic activities in several important cellular signaling pathways and molecular targets (20, 21). This finding implies that carotenoid metabolites may have greater biologic roles than their parent compounds in human health and disease. With the mapping of the human genome and the development of “-omics,” greater understanding of carotenoid metabolism and activity is expected to provide new insight into the biologic functions of carotenoids.
CHEMICAL PROPERTIES
More than 750 carotenoids with characterized chemical structures and key analytic data are listed in the Carotenoids Handbook (22), and new carotenoids continue to be identified. Among the carotenoids found in nature, approximately 40 to 50 occur in the human food chain, and 24 have been detected in human plasma and tissues (23). The most abundant carotenoids in human plasma include β-carotene, α-carotene, β-cryptoxanthin, lutein, zeaxanthin, and lycopene. These six major carotenoids account for approximately 70% of all carotenoids identified in human plasma and tissues. Carotenoids are divided into two major groups: xanthophylls, which are oxygenated carotenoids that include lutein, zeaxanthin, and β-cryptoxanthin; and carotenes, which are hydrocarbon carotenoids that are either cyclized, such as α-carotene and β-carotene, or linear, such as lycopene.
The basic structure of a carotenoid is a 40-carbon atom conjugated polyene chain, sometimes terminated by ring structures (22). The polyene chain, which typically contains a series of conjugated double bonds (e.g., -C=C-C=C-) in the central chain of the molecule, represents a chromophore that is responsible for the characteristic colors associated with carotenoids. It also causes instability in the carotenoids by making them susceptible to cleavage by oxidation, heat, light, acid, and isomerization from trans to cis forms. Conjugated systems with alternating single and double bonds result in a general delocalization of electrons across adjacent atoms that allows for resonance-stabilized structures. These structures, which impart both a capacity to act as biologic antioxidants and the ability to absorb and give off certain wavelengths of light, cause a compound to appear colored.
Fig. 31.2. Proposed metabolic pathway and chemical structures of the major nonprovitamin A carotenoids (lutein, zeaxanthin, and lycopene) found in human plasma and tissues. Cleavage of xanthophylls (lutein and zeaxanthin) by β-carotene-9′,10′-oxygenase (BCO2) may occur at either the 9,10 or 9′,10′ double bond to produce 3-OH-β-apo-10′-carotenal and β-ionone, or 3-OH-β-ionone and β-apo-10′-carotenal, respectively. Cleavage of cis-lycopene by BCO2 may occur at either the 9,10 or 9′,10′ double bond to produce apo-10′-lycopenal, which can be oxidized to apo-10′-lycopenoic acid or reduced to apo-10′-lycopenol. ADH, alcohol dehydrogenase; ALDH, aldehyde dehydrogenase.
β-Carotene, α-carotene, and β-cryptoxanthin are important sources of vitamin A (see the chapter on vitamin A). All provitamin A carotenoids have one or two β-ionone rings (see Fig. 31.1). β-Carotene has a symmetric chemical structure characterized by a long carbon chain with alternating double and single bonds, terminated at each end by a ring structure (see Fig. 31.1). Lutein and zeaxanthin both have oxygen added to their ionone ring, but each differs from the other in the position of the double bond in one of the ionone rings (see Fig. 31.2). Lycopene has a symmetric open chain structure without a ring (see Fig. 31.2).
Major chemical data for these carotenoids are found in the Carotenoids Handbook (22). Carotenoids usually occur in nature as all-trans-isomers, but the known exceptions include 9-cis-β-carotene in the alga Dunaliella and the 15-cis-phytoene present in tomatoes and other organisms. The cis-trans isomerism of carbon-carbon double bonds is an important feature of the stereochemistry of carotenoids because these geometric isomers may have different biologic properties. Approximately 370 of the naturally occurring carotenoids are chiral, bearing 1 to 5 asymmetric carbon atoms; most individual carotenoids occur in nature in only a single configuration.
The term apocarotenoids or apolycopenoids refers to carotenoid derivatives in which the carbon skeleton is shortened by the removal of fragments from one or both ends of the carotenoid with the position of the point of cleavage indicated (e.g., β-apo-10′-carotenal from β-carotene [see Fig. 31.1] or apo-10′-lycopenal from lycopene [see Fig. 31.2]). The same applies to their further metabolites, such as alcohol and acid forms. Apocarotenoids are bioactive mediators in plants, in which they act as visual or volatile signals to attract pollinating and seed dispersing agents. These substances are key players in allelopathic interactions, plant defense, and plant architecture (24). Abscisic acid, formed by specific oxidative cleavage of the 11,12 carbon-carbon double bond of 9′-(Z)-neoxanthin, acts as a hormone in plants. In humans, the most important apocarotenoids are retinal and its derivatives.
DIETARY SOURCES
The major sources of carotenoids in the human diet are deeply pigmented yellow to red fruits and vegetables (Table 31.1). Common examples of carotenoid coloration in the human diet include yellow vegetables, such as corn, and egg yolk. Both are rich in carotene and lutein. β-Carotene is responsible for the orange color of carrots, lycopene for the red color of tomatoes and watermelon, and zeaxanthin for the pink color of salmon. In the green leaves of vegetables, the colors of the carotenoids are often masked by chlorophyll. A more extensive list of dietary sources of carotenoids is found in Table 31.1. Since the 1990s, great effort has been exerted on metabolic engineering of carotenoids into agricultural crops, such as golden rice, high-β-carotene maize, cassava roots, and potatoes. Currently, scientists are evaluating these new food products for their bioavailability to provide vitamin A and are assessing the potential of these products to combat vitamin A deficiency safely worldwide.
TABLE 31.1 DIETARY INTAKE, SERUM CONCENTRATIONS, AND NATURAL SOURCES OF SIX MAJOR CAROTENOIDS
CAROTENOIDS
FORMULA
MEDIAN DIETARY INTAKE (µg/d) REFERENCE RANGE (10TH-90TH PERCENTILE)
MEDIAN SERUM CONCENTRATIONS (µg/dL) REFERENCE RANGE (10TH-90TH PERCENTILE)
MAJOR NATURAL SOURCES
β-Carotene
C40H56
1,665 (774-3,580)
14.7 (6.4-35.1)
Carrots, sweet potato, spinach, broccoli, kale, pumpkin, mango, apricot, and many yellow-orange fruits and green leafy vegetables
α-Carotene
C40H56
36 (2-1.184)
3.4 (1.3-9.2)
Carrots, pumpkin, some yellow-orange fruits and green vegetables
β-Cryptoxanthin
C40H56O
88 (24-319)
8.0 (4.0-16.4)
Tangerine, orange, sweet red pepper, peach, papaya; in small amounts, in yellow-orange fruits and in maize
Lutein + zeaxanthin
C40H56O2
1,466 (714-3,021)
18.9 (11.1-33.0)
Egg yolk, broccoli, spinach, yellow pepper, kale, sweet corn, pumpkin, and all green fruits and vegetables
Lycopene
C40H56
8,031 (3,580-16,833)
22.4 (11.9-36.1)
Tomato and tomato products (e.g., sauce, pasta, soup, and juice) and watermelon
Data from the Third National Health and Nutrition Examination Survey (NHANES III), from 1988 to 1994, as discussed in Trumbo P, Yates AA, Schlicker S et al. Dietary reference intakes: vitamin A, vitamin K, arsenic, boron, chromium, copper, iodine, iron, manganese, molybdenum, nickel, silicon, vanadium, and zinc. J Am Diet Assoc 2001;101:294-301; and Monsen ER. Dietary reference intakes for the antioxidant nutrients: vitamin C, vitamin E, selenium, and carotenoids. J Am Diet Assoc 2000;100:637-40, with permission.
DIETARY INTAKE AND SERUM CONCENTRATIONS
Information on the dietary intake and serum concentrations of carotenoids (see Table 31.1) was obtained from the Third National Health and Nutrition Examination Survey (NHANES III) (25, 26). Carotenoids are not classified as essential nutrients, so values for their recommended daily intake have not been established. β-Carotene and other provitamin A carotenoids are confirmed important sources of vitamin A, but no specific recommendation exists on the percentage of total vitamin A that should be obtained from retinyl esters or carotenoids. At present, the reported values for the conversion of β-carotene to vitamin A (i.e., the number of β-carotene molecules that are nutritionally equivalent to one molecule of vitamin A) exhibit a wide range, from 2:1 for synthetic pure β-carotene in oil to 27:1 for β-carotene from vegetables (27). Several factors affect β-carotene bioavailability and conversion to vitamin A, such as the food matrix (e.g., vegetables, fruits) and the host nutritional and health status (e.g., vitamin A status, malnutrition, parasitic infection). The Institute of Medicine in the United States has not established a recommended dietary allowance or an adequate intake for β-carotene or total carotenoids. In 2003, however, the Expert Group on Vitamins and Minerals in the United Kingdom established a safe upper level of 7 mg for the daily intake of β-carotene in dietary supplements (28).
Studies show that common nonsynonymous single nucleotide polymorphisms (SNPs) exist in the human β-carotene cleavage enzyme gene (BCO1) and occur at high frequencies, thus altering β-carotene metabolism (29). These studies may provide an explanation for the various phenotypes observed in β-carotene absorption and metabolism. In addition, the studies indicate that genetic variability in the population should be taken into account for future recommendations of vitamin A supplementation.
ANALYSIS
High-performance liquid chromatography (HPLC) is a powerful means of analyzing carotenoid composition and determining concentrations after the extraction of carotenoids from plasma, tissue, and food samples. The intrinsic ultraviolet (UV)-visible absorption spectrum provides the first criterion for identifying a carotenoid and is the basis of quantitative analysis. Carotenoids have a characteristic absorption spectrum, and their concentrations can be calculated from specific extinction coefficients.
The chemical data on major carotenoids, including UV-visible spectra and extinction coefficients, have been described by Britton et al (22). The photodiode array detector allows simultaneous monitoring at a range of selected wavelengths and provides UV/Vis spectra online for each component of a chromatogram as an aid to identification. Linked HPLC-mass spectrometry and HPLC-nuclear magnetic resonance (NMR) techniques are becoming more readily available.
The identification of carotenoids as known compounds should, at the minimum, be based on the following: (a) the UV-visible absorption spectrum (λmax) must be identical to that of an authentic sample; (b) the chromatographic properties must be identical to those of an authentic sample in HPLC, and cochromatography with an authentic sample should be demonstrated; and (c) if possible, a mass spectrum should be obtained that allows for at least one confirmation of the molecular mass. For quantitative analysis by HPLC, an internal control, such as echinenone, is needed to assess the efficiency of extraction procedures. Full elucidation of the structure requires a fully assigned NMR spectrum and, for chiral compounds, comparison of a circular dichroism spectrum with that of an authentic reference sample.
Carotenoids are unstable and are vulnerable when exposed to oxygen, heat, light, and acid. Precautions and special procedures must be used to minimize the risk of degradation and the formation of artifacts. All analytic procedures should be carried out in an inert atmosphere (nitrogen or argon), at room temperature (˜20°C), in the dark or in diffused light, under acid-free conditions, and with freshly purified, peroxide-free solvents. Geometric (cis-trans) isomerization occurs readily when carotenoids are exposed to factors such as light or heat and slowly occurs even in isolated or purified samples. Plasma or tissue samples should be stored at -80°C to minimize degradative reactions and isomerization.
Noninvasive resonance Raman spectroscopic techniques show promise for measuring carotenoids in situ in the skin and the retina (30, 31). Carotenoids in the skin of the palm of the hand can be measured using a portable Raman device. Emission of light of 488 nm is used to estimate total carotenoids, and emission of light of 514 nm is used to estimate lycopene. The lack of power to separate individual carotenoids, except lycopene, is a limit to research, but great potential exists for monitoring carotenoid status from fruit and vegetable intake or the effect of carotenoid supplementation in human trials. Lutein and zeaxanthin, major macular pigments of the human retina, can also be measured using resonance Raman spectroscopic techniques (30). This non-invasive detection technique can be a screening method for macular pigment levels in the general population.
Stable isotope techniques that use intrinsically labeled carotenoids have proven useful in determining carotenoid bioavailability, bioconversion, and bioefficacy from different human food sources (27). Although these methods are costly and complex, they can distinguish between dosed and endogenous carotenoids and can determine the vitamin A equivalency of provitamin A carotenoids.
ABSORPTION, BIOAVAILABILITY, AND TRANSPORT
Much of the research on carotenoids to date has concentrated on β-carotene. The efficiency of absorption of a moderate dose of β-carotene in oil is approximately 9% to 22%. Humans (along with monkeys, ferrets, and gerbils, but excluding rats, mice, and rabbits unless they are given very high doses) absorb a significant portion of intact carotenoids directly and circulate or accumulate them in their plasma, liver, and peripheral tissues. The median concentrations of carotenoids were reported in NHANES III (see Table 31.1). The half-life of plasma carotenoids ranges from up to 12 days for β-carotene, α-carotene, and cryptoxanthin, to 12 to 33 days for lycopene, and 33 to 61 days for zeaxanthin and lutein (32).
The bioavailability of β-carotene from vegetables is generally low (33). The mnemonic acronym SLAMENGHI lists the major contributors that affect carotenoid bioavailability. SLAMENGHI stands for species of carotenoids, linkages molecular level, amount of carotenoid, matrix effectors, nutrient status, genetics, host-related factors, and interactions among these variables (33). These factors are discussed in detailed in the literature (34, 35).
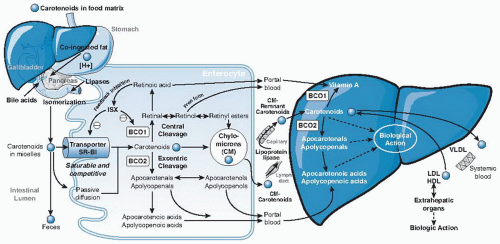
Carotenoids embedded in their food matrix cannot be absorbed efficiently. Food processing and cooking that cause the mechanical breakdown of the food matrix and the release of carotenoids can improve intestinal absorption. After release from the food matrix, ingested carotenoids must be emulsified and solubilized into micelles before they are absorbed into the intestinal mucosa (Fig. 31.3).
Previous research assumed that the process of carotenoid absorption occurred by passive diffusion. More recent studies, however, indicated the involvement of an active process for the uptake of carotenoids through the scavenger receptor class B type 1 protein transporter (SR-B1) (36). SR-B1 is found in the human small intestine, as well as the liver, adrenals, ovaries, placenta, kidneys, prostate, and brain. Therefore, SR-B1 may be partially responsible for the transport of carotenoids from lipoprotein to tissues and from tissues to lipoproteins (37). A diet-responsive regulatory network involving the intestinespecific homeobox (ISX) transcription factor was shown to regulate intestinal β-carotene uptake and vitamin A production by a negative feedback regulatory mechanism (36). ISX repressed both expression of intestinal BCO1 (38) and SR-B1 (39), which facilitates the absorption of dietary lipids and carotenoids (34). Because ISX is under the control of retinoic acid and retinoic acid receptor (RAR) receptor-dependent mechanisms, during vitamin A insufficiency, both BCO1 expression and SR-B1 expression are induced to increase absorption and conversion of β-carotene to vitamin A (see Fig. 31.3). Cleavage of β-carotene by BCO1 produces retinal, which can be oxidized to retinoic acid. Retinoic acid induces the expression of the ISX transcription factor and then represses expression of both BCO1 and SR-B1, to complete the dietary feedback mechanism (see Fig. 31.3).
Another protein, CD36, a surface membrane glycoprotein in the duodenum and jejunum involved in the uptake of long-chain fatty acids and oxidized low-density lipoproteins (LDLs), may also play a role in movement of carotenoids into cells. Although the same factors that influence the absorption of β-carotene may affect other carotenoids similarly, more investigation of the absorption of individual carotenoids and their cis isomers is needed.
Fig. 31.3. Simplified schematic illustration of absorption, metabolism, and transport of carotenoids. BCO1, β-carotene-15,15′-oxygenase; BCO2, β-carotene-9′,10′-oxygenase; HDL, high-density lipoprotein; LDL, low-density lipoprotein; VLDL, very-low-density lipoprotein. (See text for detailed information).
After β-carotene is taken up by the mucosa of the small intestine, it is either cleaved by BCO1 or BCO2 into vitamin A and other metabolites or packaged into chylomicrons and secreted into the lymphatic system for transport to the liver and other peripheral tissues (see Fig. 31.3). Some polar metabolites can be directly transported into the liver through the portal blood system (40). β-Carotene, retinyl esters, retinol, and the less polar metabolites are absorbed into the lymph, whereas the more polar metabolites, which include β-apocarotenals, retinoyl-β-glucuronide, retinyl-β-glucuronide, and retinoic acid, are taken up directly into the portal blood (40). The differential absorption of β-carotene and its metabolites into lymph or portal blood seems to depend on the polarity of the metabolites involved.
Only gold members can continue reading. Log In or Register to continue